|
||||
|
ГЛАВА 3 Нервный импульс, яды и противоядия Все большее распространение получают теперь химические соединения, избирательно воздействующие на функции нервной системы. Это связано, во-первых, с постоянно увеличивающимися возможностями контакта современного человека с большим числом нейротоксических веществ в промышленности и сельском хозяйстве (препараты группы гидразина, ФОС, карбаматы и др.) и, во-вторых, с расширением диапазона применения средств предупреждения и лечения различных нервно-психических расстройств (транквилизаторы, антидепрессанты, наркотики, аналептики и др.). Достоверные научные данные дают основание объединить названные группы веществ одним общим фармакологическим свойством — способностью влиять на передачу нервного импульса. Наиболее демонстративно это иллюстрируется рассмотрением биохимических аспектов действия ФОС и соответствующих антидотов. Фосфорорганические соединения Синтезированные впервые в конце прошлого века ФОС привлекли к себе особенно пристальное внимание химиков и токсикологов с середины 30-х годов, когда их свойства более тщательно были исследованы из-за неожиданно обнаруженной высокой токсичности. Именно тогда в одной из лабораторий германской фирмы «И. Г. Фарбениндустри» под руководством Шрадера были синтезированы фосфорорганические инсектициды, проявлявшие биологическую активность в очень малых дозах. В дальнейшем в связи с подготовкой фашистской Германии к химической войне эта лаборатория переключилась на работы по созданию высокотоксичных ФОС, предназначенных для военных целей. Там были синтезированы такие известные отравляющие вещества из этого класса, как табун, зарин, зоман. Распространение ФОС обусловлено прежде всего повсеместным их использованием в качестве ядохимикатов. Достаточно назвать в связи с этим такие инсектициды, как хлорофос, фосфамид, карбофос, октаметил. Возрастает и число фосфорорганических медикаментозных средств, используемых в невропатологии, офтальмологии, хирургии (армии, фосфакол, фосарбид и др.). Множество ФОС применяется в химической промышленности, в частности, в качестве исходных и промежуточных продуктов органического синтеза. В нашей стране большие заслуги в синтезе и разработке способов применения ФОС, имеющих народнохозяйственное значение, принадлежат казанской школе химиков-органиков, возглавляемой академиком Б. А. Арбузовым. Обширные исследования были проведены этими учеными в области превращения эфиров кислот трехвалентного фосфора в эфиры фосфоновых кислот. Советскими и зарубежными исследователями были раскрыты и многие другие химические свойства этих веществ. Однако, как справедливо отмечает Франке (ГДР), «развитие химии ФОС отнюдь не завершено. Среди этих соединений можно ожидать появления еще более токсичных веществ. Большие возможности комбинаций, заметное изменение свойств при небольших изменениях строения создают богатое поле деятельности для химиков, фармацевтов, врачей, биологов, физиологов и др.».[69] По своему химическому строению ФОС представляют собою эфиры кислот пятивалентного фосфора (фосфорной, тнофосфорной, фосфоновой и др.). Их общий вид хорошо иллюстрируется следующей структурной формулой: 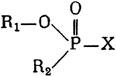 где P — атом фосфора, R1 и R2, как правило, органические радикалы, а X — галоген (Cl, F), или CN-гpуппa, или производное бензола (например, нитрофенол). Будучи в основном малолетучими жидкостями, ФОС способны проникать во внутренние среды организма через неповрежденную кожу и слизистые оболочки вследствие высокой липидотропности. Источником отравлений могут быть зараженная пища и вода, а также воздух, содержащий пары и аэрозоли ФОС. Однако картина отравления мало зависит от путей поступления ФОС в организм и в основном сводится к нарушениям функции цейтральной нервной системы, а также мышечной, дыхательной, сердечно-сосудистой систем, желудочно-кишечного тракта и органа зрения. У тяжело отравленного быстро появляется беспокойство, чувство страха, возбуждение, судороги. Эти симптомы развиваются на фона приступов удушья и кашля из-за спазма бронхов и обильного выделения секрета бронхиальных желез. Появляются боли в области сердца, расстраивается его ритм, подъем кровяного давления сменяется падением. Возникают боли в животе, спазмируется кишечник, наблюдается понос. Очень характерны изменения со стороны глаз: резко суживается зрачок, появляется слезотечение, нарушается функция зрения, особенно аккомодация, т. е. способность настраивать глаз на ближнее или дальнее видение. При попадании в организм смертельных доз ФОС и несвоевременном или нерациональном лечении быстро развивается обморочное состояние, наступает паралич дыхательного центра, что и является чаще всего непосредственной причиной гибели отравленных. Холиеэстераза и ацетилхолин Согласно существующим представлениям, в основе механизма действия ФОС лежит избирательное торможение ими фермента ацетилхолинэстеразы, или просто холинэстеразы, которая катализирует гидролиз ацетилхолина-химического передатчика (медиатора) нервного возбуждения. Различают 2 типа холинэстеразы: истинную, содержащуюся преимущественно в тканях нервной системы, в скелетной мускулатуре, а также в эритроцитах, и ложную, содержащуюся главным образом в плазме крови, печени и некоторых других органах. Собственно ацетилхолинэстеразой является истинная, или специфическая, холинэстераза, так как только она гидролизует названный медиатор. И именно ее в дальнейшем мы будем обозначать термином «холинэстераза». Поскольку фермент и медиатор являются необходимыми химическими компонентами передачи нервных импульсов в синапсах — контактах между двумя нейронами или окончаниями нейрона и рецепторной клеткой, следует более подробно остановиться на их биохимической роли. Ацетилхолин синтезируется из спирта холина и ацетилкоэнзима А[70] под влиянием фермента холинацетилазы в митохондриях нервных клеток и накапливается в окончаниях их отростков в виде пузырьков диаметром около 50 нм. Предполагается, что каждый такой пузырек содержит несколько тысяч молекул ацетилхолина. При этом в настоящее время принято различать ацетилхолин, готовый к секреции и расположенный в непосредственной близости от активной зоны, и ацетилхолин вне активной зоны, находящийся в состоянии равновесия с первым и не готовый к выделению в сипаптическую щель. Кроме того, имеется еще так называемый стабильный фонд ацетилхолина (до 15%), не освобождающегося даже в условиях блокады его синтеза.[71] Под воздействием нервного возбуждения и ионов Са2+ молекулы ацетилхолина переходят в синаптическую щель — пространство шириною 20–50 нм, отделяющее окончание нервного волокна (пресинаптическую мембрану) от иннервируемой клетки. На поверхности последней расположена постсинаптическая мембрана с холинорецепторами — специфическими белковыми структурами, способными взаимодействовать с ацетилхолином. Воздействие медиатора на холинорецептор приводит к деполяризации (снижению заряда), временному изменению проницаемости постсинаптической мембраны для положительно заряженных ионов Na+ и проникновению их внутрь клетки, что в свою очередь выравнивает потенциал напряжения на ее поверхности (оболочке).[72] Это дает начало новому импульсу в нейроне следующей ступени или вызывает деятельность клеток того или иного органа: мышцы, железы и др. (рис 5). Фармакологические исследования выявили существенную разницу в свойствах холинорецепторов различных синапсов. Рецепторы одной группы, проявляющие избирательную чувствительность к мускарину (яду гриба мухомора), названы мускариночувствительными, или М-холинорецепторами; они представлены главным образом в гладких мышцах глаз, бронхов, желудочно-кишечного тракта, в клетках потовых и пищеварительных желез, в сердечной мышце. Холинорецепторы второй группы возбуждаются малыми дозами никотина и поэтому названы никотино-чувствительными, или Н-холинорецепторами. К ним относятся рецепторы вегетативных ганглиев, скелетных мышц, мозгового слоя надпочечниковых желез, центральной нервной системы. 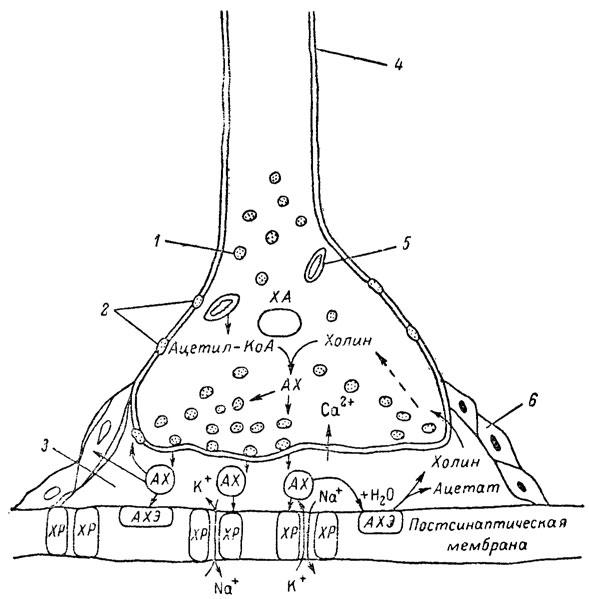 Рис. 5. Основные структурные элементы и схема функционирования холинергического синапса (Голиков, Фишзон-Рысс, 1978). 1 — синаптические пузырьки; 2 — пресинаптические центры связывания ацетилхолина; 3 — синаптическая щель; 4 — миелиновая оболочка; 5 — митохондрии; 6 — шванновские клетки; ХР — холинорецептор; АХ — ацетил-холин; АХЭ — ацетилхолинэстераза; ХА — холинацетилаза Рис. 5. Основные структурные элементы и схема функционирования холинергического синапса (Голиков, Фишзон-Рысс, 1978). 1 — синаптические пузырьки; 2 — пресинаптические центры связывания ацетилхолина; 3 — синаптическая щель; 4 — миелиновая оболочка; 5 — митохондрии; 6 — шванновские клетки; ХР — холинорецептор; АХ — ацетил-холин; АХЭ — ацетилхолинэстераза; ХА — холинацетилаза Молекулы ацетилхолина, выполнившие свою медиаторную функцию, должны быть немедленно инактивированы, в противном случае будет нарушена дискретность в проведении нервного импульса и проявится избыточная функция холинорецептора. Именно это осуществляет холинэстераза, мгновенно гидролизующая ацетилхолин. Каталитическая активность холинэстеразы превышает почти все известные ферменты: по разным данным, время расщепления одной молекулы ацетилхолина составляет около одной миллисекунды, что соизмеримо со скоростью передачи нервного импульса. Осуществление столь мощного каталитического эффекта обеспечивается наличием в молекуле холинэстеразы определенных участков (активных центров), обладающих исключительно хорошо выраженной реакционной способностью по отношению к ацетилхолину.[73] Будучи простым белком (протеином), состоящим только из одних аминокислот, молекула холинэстеразы, как теперь выяснено, исходя из ее молекулярной массы, содержит от 30 до 50 таких активных центров. 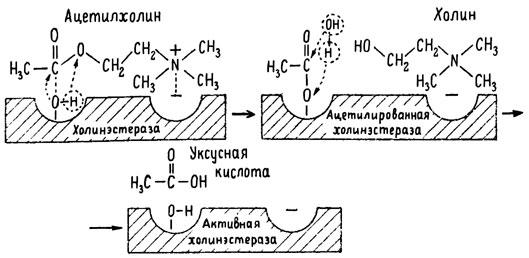 Рис. 6. Ферментативный гидролиз ацетилхолина. Объяснение в тексте Рис. 6. Ферментативный гидролиз ацетилхолина. Объяснение в тексте Как видно из рис. 6, участок поверхности холинэстеразы, непосредственно контактирующий с каждой молекулой медиатора, включает 2 центра, расположенных на расстоянии 0,4–0,5 мм: анионный, несущий отрицательный заряд, и эстеразный. Каждый из этих центров образован определенными группами атомов аминокислот, составляющих структуру фермента (гидроксилом, карбоксилом и др.). Ацетилхолин благодаря положительно заряженному атому азота (так называемой катионной головки) ориентируется за счет электростатических сил на поверхности холинэстеразы. При этом расстояние между атомом азота и кислотной группой медиатора соответствует расстоянию между активными центрами фермента. Анионный центр притягивает к себе катионную головку ацетилхолина и тем самым способствует сближению его эфирной группировки с эстеразным центром фермента. Затем рвется эфирная связь, ацетилхолин разделяется на 2 части: холиновую и уксусную, остаток уксусной кислоты присоединяется к эстеразному центру фермента и образуется так называемая ацетилирозанная холинэстераза. Этот крайне непрочный комплекс мгновенно подвергается спонтанному гидролизу, что освобождает фермент от остатка медиатора и приводит к образованию уксусной кислоты. С данного момента холииэстераза снова способна выполнять каталитическую функцию, а холин и уксусная кислота становятся исходными продуктами синтеза новых молекул ацетилхолина. Механизм действия фосфорорганических соединений Многие ФОС вследствие особого химического сродства к холинэстеразе ингибируют, т. е. блокируют, ее молекулы посредством взаимодействия с эстеразным центром. Как это видно из приводимой схемы, молекула фосфорор-ганического яда реагирует с гидроксильной группой фермента, причем анионный его центр участия в реакция не принимает: 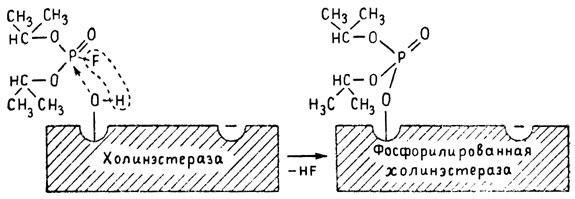 Однако в начале 50-х годов в Швеции в лаборатории Таммелина были синтезированы такие ФОС, которые реагируют и с анионным центром фермента. При этом химики исходили из предположения, что если яд будет сходен по структуре с естественным субстратом холинэстеразы (ацетилхолином), то он окажет на этот фермент более сильное ингибирующее действие. И действительно, такими мощными антихолинэстеразными веществами оказались соединения, которые содержат в своем составе остаток холина, как например метилфторфосфорилхолин. Совершенно естественно, что при его взаимодействии с холинэстеразой положительно заряженный атом азота будет реагировать с анионным центром фермента. Это обеспечивает дополнительный контакт яда с активной поверхностью холинэстеразы и делает связь между ними более прочной: 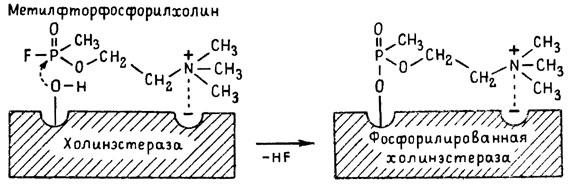 Можно представить также одномоментное ингибирование одной молекулой метилфторфосфорилхолнна двух молекул фермента: одну по анионному, вторую — по эстеразному центру. Как бы то ни было, яды Таммелина оказались в десятки раз более токсичными, чем даже такое сильнодействующее ФОС, как зарин. Образовавшаяся фосфорилированная холинэстераза в отличие от ацетилированной является соединением достаточно прочным и не подвергается самопроизвольному гидролизу. Оказалось, что процесс ингибирования холинэстеразы является двухэтапным. Вначале, на первом этапе, происходит обратимое, т. е. непрочное, ее блокирование, и лишь на втором этапе наступает необратимое блокирование фермента. Оба эти этапа являются результатом сложной, до конца еще не выясненной молекулярной перестройка в комплексе «ФОС-холинэстераза». Забегая несколько вперед, отметим значимость данного явления для практики применения некоторых антидотов, действие которых заключается в разрыве химической связи между ядом и ферментом. Таким образом, под влиянием антихолинэсте — разных веществ тормозится разрушение молекул ацетилхолина и он продолжает оказывать непрерывное действие на холинорецепторы. Отсюда следует, что отравление ФОС есть не что иное, как генерализованное перевозбуждение холинорецепторов, вызванное интоксикацией эндогенным, т. е. имеющим внутреннее происхождение, ацетилхолином. Вот почему основные симптомы отравления ФОС можно трактовать как проявление избыточной, нецелесообразной для организма деятельности ряда структур и органов, которая обеспечивается ацетилхолиновой медиацией (прежде всего это — функция нервных клеток, поперечнополосатых и гладких мышц, различных желез). В настоящее время имеются доказательства непосредственного возбуждающего влияния некоторых ФОС на холинорецепторы. Тем самым не исключается, что ФОС оказывают токсический эффект, минуя холинэстеразный механизм: 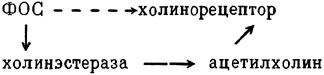 В последние 10–15 лет токсикологи все чаще обращают внимание на эту особенность механизма влияния ФОС на биоструктуры. В частности, такое их неантихолинэстеразное действие проявляется в прямом возбуждении Н-холинорецепторов, от чего, как показывают экспериментальные данные, в свою очередь зависят никоти — ноподобные эффекты ФОС. В то же время их мускарино — подобное действие с достаточным основанием теперь рассматривается как результат ингибирования холинэстеразы. Что касается сравнительной выраженности мускариновых и никотиновых эффектов у различных антихолинэстеразных веществ, то, согласно исследованию В. Б. Прозоровского,[74] можно рассматривать 3 их группы: 1) вызывающие преимущественное возбуждение М-холинорецепторов (эзерин, нибуфин, хлорофос); 2) вызывающие возбуждение как М-, так и Н-холинорецепторов (фосфакол, армин, диизопропилфторфосфат); 3) вызывающие преимущественное воздействие на Н-холинорецепторы (прозерин, тиофос, меркаптофос). Из изложенного следует, по крайней мере теоретически, что при отравлении антихолинэстеразными ядами, в том числе ФОС, антидотами могли бы быть: 1) вещества, вступающие в прямое химическое взаимодействие с ядами; 2) вещества, тормозящие синтез и выход ацетилхолина в синаптическую щель; 3) вещества, замещающие поврежденный ядами фермент (т. е. препараты холинэстеразы); 4) вещества, препятствующие контакту яда с ферментом и тем самым защищающие его от токсического воздействия; 5) вещества, препятствующие контакту ацетилхолина с холинорецептором; 6) вещества, восстанавливающие активность фермента посредством вытеснения яда с его поверхности (т. е. реактивирующие структуру холинэстеразы). Многочисленные токсикологические эксперименты показали, что всем этим веществам присуща та или иная степень специфического воздействия на токсический процесс, однако наибольшее практическое значение имеют 2 последние группы противоядий. Рассмотрим подробнее механизмы их действия. Атропин и атропиноподобные вещества 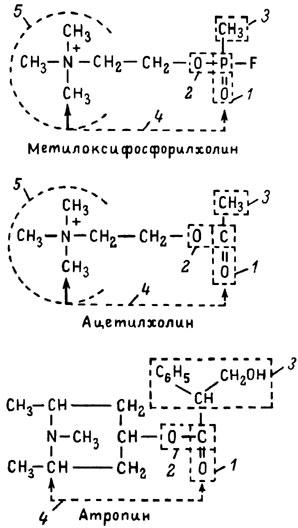 Рис. 7. Химическое строение молекулы высокотоксичного ФОС, ацетилхолина и атропина (Gille, 1971). 1 — двойная связь (С=0 или Р=0); 2 — эфирная связь; 3 — алкильная группа у Р- или С-атома; 4 — одинаковое расстояние между центрами действия; 5 — катионная головка молекулы Рис. 7. Химическое строение молекулы высокотоксичного ФОС, ацетилхолина и атропина (Gille, 1971). 1 — двойная связь (С=0 или Р=0); 2 — эфирная связь; 3 — алкильная группа у Р- или С-атома; 4 — одинаковое расстояние между центрами действия; 5 — катионная головка молекулы Антидотный механизм, о котором пойдет речь, состоит в блокировании молекулами противоядий холинорецепторов и в прерывании или тормошении вследствие этого медиаторной функции ацетилхолина. Вещества такого типа действия получили в фармакологии название холиполитиков. Помимо уже знакомого нам атропина в настоящее время находят применение амизил, амедин, бензацин, апрофен, пентафен, тропацин, спазмолитин, мепанит и другие препараты, часть из которых значительно превосходит атропин по силе действия. К тому же атропин блокирует преимущественно М-холинорецепторы и мало влияет на никотиноподобное действие ФОС (т. е. слабо устраняет нарушения в передаче импульса с нерва на мышцу, деятельности сердца, надпочечниковых желез и некоторые другие симптомы). В то же время такие холинолитики, как пентафен, циклодол, динезин, а также ганглиоблокаторы[75] — гексоний, пентамин и др., проявляя антимускариновое действие, одновременно ослабляют никотиноподобные эффекты ФОС. Поэтому при отравлении антихолинэстеразными веществами наиболее рациональным считается комбинированное применение различных холинолитиков. Так, в одной из работ профессора Ю. С. Кагана[76] была показана высокая эффективность комбинации пентафена и мепанита при отравлении животных меркаптофосом — высокотоксичным ингибитором холинэстеразы. Автор учитывал при этом данные, полученные в лаборатории профессора М. Я. Михельсона, о неодинаковом действии на те или иные звенья нервной системы различных холинолитиков, являющихся третичными аминами (пентафен) или имеющими в своем составе четвертичный азот (мепанит). В этой связи следует иметь также в виду результаты опытов, приведенные в уже упомянутой работе В. Б. Прозоровского, который показал, что отравление антихолинэстеразными веществами, преимущественно возбуждающими М-холинорецепторы, легко предупреждается и лечится с помощью одних М-холинолитиков (например, атропином). Если же последние применять при отравлении ядами, действующими главным образом на Н-холинорецепторы, то они оказываются практически неэффективными, а антидот. ный эффект достигается только при использовании Н-холинолитиков (например, пентафена). Эти интересные данные, подтверждающие высокую специфичность холинолитических противоядий, дополняются установленной необходимостью обязательного комбинирования М-холинолитиков с Н-холинолитиками для успешного лечения отравлений веществами, возбуждающими как М-, так и Н-холинорецепторы. В чем же состоит молекулярный механизм действия холинолитических антидотов? Прежде всего надо отметить структурное сходство холинолитиков, в том числе атропина, с ацетилхолином и некоторыми ФОС (рис. 7). Данное обстоятельство, по-видимому, в немалой степени определяет химическое сродство этих противоядий и ряда ФОС к холинорецептору, а также существование конкурентного антагонизма между ними и медиатором за связь с рецептором. При этом в определенных дозах холинолитик проявляет большую способность взаимодействовать с холинорецептором, чем медиатор, и даже может вытеснять последний с холинорецептора. Разумеется, такой антидот будет блокировать и нормально функционирующие холинорецепторы, свободные от избытка ацетилхолина. Вот почему антидоты типа атропина могут применяться не только с лечебной, но и с профилактической целью: предварительное введение животным 1 мл 0,1%-ного водного раствора сульфата атропина (1 мг) внутримышечно или подкожно предотвращает или резко ослабляет отравление антихолинэстеразными веществами. 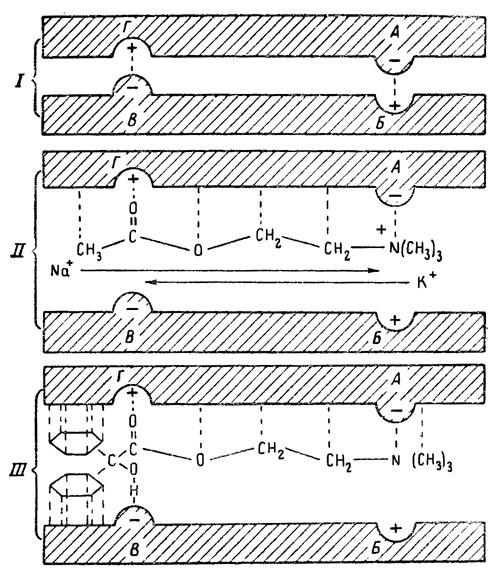 Рис. 8. Схематичное изображение активного участка холинорецептора (ХР), взаимодействия ацетилхолина с ХР и взаимодействия холинолитического антидота с ХР (Кузнецов, Голиков, 1962). Рецепторная часть ХР (I) построена из двух белковых молекул, на поверх ностях которых существует 2 активных центра, В одном из них — анионном — белковые цепи связаны ионными связями (А---Б), в другом — эстерофильном — водородными (В---Г). Молекула ацетилхолина, взаимодействуя с рецептором (II), присоединяется своим катионным концом к анионному центру, разрывая связь А---Б. Эфирная группировка ацетилхолина соединяется с эстерофильным центром, разрывая водородную связь В---Г В результате в структуре рецепторного белка возникает разрыв, образуется коридор, по которому осуществляется уравнивание концентраций ионов К+ и Na+ снаружи и внутри клетки, нервный импульс проходит через синапс. Молекула холинолитика (III) выключает ХР из механизма нервной передачи Рис. 8. Схематичное изображение активного участка холинорецептора (ХР), взаимодействия ацетилхолина с ХР и взаимодействия холинолитического антидота с ХР (Кузнецов, Голиков, 1962). Рецепторная часть ХР (I) построена из двух белковых молекул, на поверх ностях которых существует 2 активных центра, В одном из них — анионном — белковые цепи связаны ионными связями (А---Б), в другом — эстерофильном — водородными (В---Г). Молекула ацетилхолина, взаимодействуя с рецептором (II), присоединяется своим катионным концом к анионному центру, разрывая связь А---Б. Эфирная группировка ацетилхолина соединяется с эстерофильным центром, разрывая водородную связь В---Г В результате в структуре рецепторного белка возникает разрыв, образуется коридор, по которому осуществляется уравнивание концентраций ионов К+ и Na+ снаружи и внутри клетки, нервный импульс проходит через синапс. Молекула холинолитика (III) выключает ХР из механизма нервной передачи Молекулярная сущность холинолитического антидотного эффекта иллюстрируется рис. 8, где в качестве холинолитика взят один из синтетических атропиноподобных препаратов и приведена вероятная схема его взаимодействия с холинорецептором. В результате такого взаимодейетвия обе белковые цепи холинорецептора как бы сшиваются и проведение нервного импульса становится невозможным.[77] Таким образом, холинолитические антидоты, образуя с холинорецептором комплекс, недоступный для ацетилхолина, на более или менее длительный срок выключают рецептор из механизма нервной передачи, что и определяет в конечном счете их действенность при отравлениях антихолинэстеразными ядами. Важно иметь в виду, что успешное лечение развившейся интоксикации ФОС достигается лишь высокими дозами атропина и других холинолитиков, причем в тяжелых случаях их эффективная разовая доза может превысить в несколько раз установленную суточную терапевтическзую дозу. Это связано с тем, что на фоне избыточной функции холинорецепторов чувствительность их к холинолитикам резко снижается. Известны случаи тяжелых отравлений ФОС, для лечения которых было израсходовано несколько сотен миллиграммов атропиноподобных веществ. Кроме того, ограниченная продолжительность действия этих препаратов диктует необходимость многократного введения их в организм при острых отравлениях. Но здесь тяжело отравленного поджидает опасность переатропинизации, признаки которой (сухость и покраснение кожи и слизистых оболочек, сердцебиение, шум в ушах, галлюцинации, расширение зрачков) указывают па необходимость немедленного прекращения антидотной терапии. Американский центр по лечению отравлений рекомендует отменять введение холинолитиков, как только у отравленного сужение зрачка сменится его расширением. Понятно, что препараты, снижающие силу и продолжительность действия холинолитиков при их длительном применении в массивных дозах, будут предотвращать избыточное угнетение холинорецепторов. Одним из ниx оказался поливинол — синтетический коллоидный заменитель плазмы, который удобно комбинировать с атропиноподобными веществами.[78] В связи с явлениями переатропинизации мы сделаем небольшое отступление. Прежде всего надо отметить, что употребление в больших дозах (количествах) настоев из растений семейства пасленовых или случайное попадание в пищу атропиноподобных веществ может привести к отравлениям, которые протекают с характерными нарушениями высшей нервной деятельности. О таких отравленных в народе говорят: «Белены объелся».[79] У них нарушается ориентировка во времени и пространстве, речь становится невнятной, возникают яркие зрительные галлюцинации, бред, расстраивается координация движений, появляется двигательное возбуждение. Вот как развиваются у людей симптомы отравления в зависимости от дозы атропина (мг):[80] • Сухость кожи, замедление пульса — 0,5 • Сухость во рту, жажда — 0,5–1 • Расширение зрачков, учащение пульса — 1–2 • Беспокойство, мышечная слабость, затруднение глотания, головная боль — 3–5 • Максимальное расширение зрачков, нарушение мышечной координации — 7 • Апатия, галлюцинация, бред, потеря сознания — 10 Нечто подобное наблюдается при попадании в организм синтетических психотомиметиков — производных гликолевой кислоты, например дитрана и бенактизина. Эти яды оказались веществами, блокирующими преимущественно М-холинорецепторы головного мозга и тем самым затрудняющими или извращающими передачу нервных импульсов в центральных холинергических структурах.[81] Совершенно очевидно, что их функциональными антагонистами могли бы оказаться вещества, способствующие стабилизации ацетилхолина в синапсах. И действительно, выраженный антидотный эффект при отравлениях холинолитическими соединениями можно наблюдать, например, при применении некоторых обратимых ингибиторов холинэстеразы, т. е. веществ, временно, нестойко угнетающих фермент. В их числе — прозерин, который взаимодействует с холинэстеразой по следующей схеме: 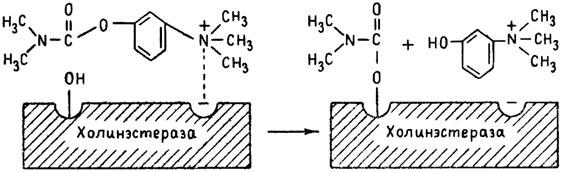 При этом молекула прозерина благодаря положительно заряженному атому азота ориентируется за счет электростатических сил на поверхности холинэстеразы. В отличие от комплекса «яд-фермент» соединение прозерина с холинэстеразой сравнительно непрочно и в течение 2–4 ч подвергается спонтанному гидролизу, что приводит к восстановлению активности фермента. Однако, вероятно из-за наличия в молекуле прозерина положительно заряженного четвертичного атома азота, он слабо проникает через гематоэнцефалический барьер и потому недостаточно взаимодействует с холинэстеразой головного мозга. Данного недостатка лишен другой антихолинэстеразный препарат — физостигмин (эзерин), растительный аналог прозерина. Этот алкалоид является действующим началом ядовитых калабарских бобов — африканского тропического растения Physostigma venenosum Balf., которое употреблялось жителями Западной Африки для производства «божьего суда» над провинившимися. Практически используется салицилат физостигмина, который при приеме внутрь в дозе около 3–4 мг оказывает антидотное действие. Закономерно предположить, что и ФОС как антихолинэстеразные вещества будут проявлять подобное действие. Поэтому нет ничего удивительного в том, что такие лекарственные препараты из группы ФОС, как фосфакол и армин, уже в малых дозах снимают симптомы интоксикации холпнолитическими ядами. И еще об одной группе антагонистов холинолитиков следует здесь упомянуть. Это вещества, возбуждающие холинорецепторы, т. е. действующие подобно ацетилхолину. К ним относятся такие синтетические препараты, как ацетилхолин-хлорид, карбахолин, а также растительный алкалоид пилокарпин. При этом в отличие от ацетилхолина антидотные эффекты карбахолина и пилокарпина проявляются в течение более длительного времени, так как данные препараты не гидролизуются под влиянием холинэстеразы. Надо иметь в виду, что в основе защитного действия веществ типа ацетилхолина лежит их конкурентный антагонизм с атропиноподобными соединениями за связь с одной и той же биохимической структурой — холинорецептором. Тем самым реализуется разнонаправленное действие двух групп веществ на одни и те же физиологические функции. Как это на первый взгляд ни парадоксально, обратимые ингибиторы холинэстеразы могут проявлять себя и как антидоты ФОС. Такое их действие основано на защите холинэстеразы от стойкого угнетения ядом. Например, предварительное введение прозерина животным снижает процент их гибели при отравлении смертельными дозами ФОС. Но введение прозерина на фоне выраженного действия яда, напротив, усиливает его токсичность. По-видимому, здесь уже сказывается потенцирование действия обратимого и необратимого ингибиторов холинэстеразы. Реактиваторы холинзстеразы Второй практически значимый антидотный механизм при отравлениях антихолинэстеразными ядами, как уже отмечалось, состоит в восстановлении (реактивировании) каталитической функции ацетилхолинэстеразы. Принцип реактивации заключается в том, что достаточно сильный реагент, каким является антидот, разрушает химическую связь между ядом и ферментом. Следовательно, антидотный эффект проявляется уже после того, как токсичное вещество инактивировало биохимическую рецептивную структуру. Наиболее ценными антидотами такого рода являются химические вещества, которых объединяет наличие в молекуле оксимной (=N-ОН) группировки и которые поэтому получили название оксимов. Из большого их числа следует прежде всего назвать те, которые находят практическое применение: дипироксим (ТМБ-4), пралидоксим (2-ПАМ), изонитрозин, токсогонин. Механизм их реактивирующего действия можно видеть на примере взаимодействия дипироксима с комплексом «ди-изопропилфторфосфат-холинэстераза»:  Как видно из приведенной схемы, наличие положительно заряженного азота в молекуле дипироксима помогает ее сближению с ингибированным ферментом.[82] При этом рвется связь яда с холинэстеразой и восстанавливается ее структура и функция. Затем реактиватор взаимодействует с ядом, образуя нетоксичный комплекс «дипироксим-ФОС», который в дальнейшем превращается в безвредные для организма продукты. Из структуры дипироксима следует, что одна его молекула может одномоментно участвовать в процессе реактивирования 2 молекул фермента. Это, по-видимому, лежит в основе высокой антидотной активности данного препарата. Но реактиваторы холинэстераз весьма эффективны также при профилактическом введении в организм, что связывается с их способностью обратимо тормозить холинэстеразу (вероятно, подобно прозерину) и тем защищать ее от последующего угнетения ФОС. Очень важно стремиться ввести отравленному реактиватор в максимально короткие сроки от момента проникновения яда в организм, так как оторвать яд от фермента практически можно лишь на этапе обратимого его ингибирования. Ясно также, что чем длиннее этот этап, тем больше шансов на успех применения реактиваторов. Для ряда сильнодействующих ФОС данное время составляет примерно 30–100 мин. При отравлении антихолинэстеразными ядами наиболее выраженным антидотным действием реактиваторы обладают тогда, когда они применяются совместно с холино-литиками. Именно такая комбинация считается самой рациональной, тем более что реактиваторы значительно усиливают антидотное действие холинолитиков. Это, например, было показано в работах зарубежного исследователя Келле[83] и у нас Г. К. Шеиным.[84] Так, Г. К. Шеин установил, что дипироксим совместно с атропином и арпеналом дает выраженный антидотный эффект при воздействии смертельных доз ФОС, в то время как сами эти холинолитики в тех же дозах не спасали животных от гибели. В последнее время появились экспериментальные доказательства свойства реактиваторов холинэстераз непосредственно реагировать с ФОС с образованием нетоксичных комплексов или необратимо трансформировать их молекулы, или же ускорять их гидролиз в организме. Это, например, можно проиллюстрировать реакцией пралидоксима с зарином:  Такое действие оксимов неизмеримо возрастает в случае их профилактического применения, что дает возможность защищать подопытных животных от смертельных доз некоторых ФОС.[85] Можно допустить, что в данном случае антидот как бы поджидает яд в организме. Ряд авторов допускают наличие у реактиваторов и других антидотных свойств. Так, не исключается, что оксимы способны взаимодействовать с холинорецепторами и даже выполнять функцию холинэстеразы, т. е. инактивировать ацетилхолин. В частности, существует мнение, что реактиватор, с одной стороны, способен защищать холинорецептор от воздействия избытка медиатора, а с другой — взаимодействовать с комплексом «ФОС-хелинорецептор», освобождая последний от яда. Все это дает основание считать реактиваторы холинэстераз антидотами многостороннего действия. В связи с изложенным небезынтересно исследование,[86] которое доказывает высокую специфичность реактиваторов холинэстераз как антидотов. Оно посвящено веществу под названием севин, представителю новой группы антихолинэстеразных ядохимикатов, производных карбаминовой кислоты ( Заканчивая описание основных антидотных механизмов при отравлениях антихолинэстеразными веществами, обратим внимание на любопытные результаты одной научной работы. Совершенно неожиданно защитное действие по отношению к ФОС выявилось у самих ФОС. Некоторое время назад академик Б. А. Арбузов с сотрудниками[87] сообщили о новых свойствах ряда ФОС с низкой токсичностью. Оказалось, что ФОС — производные кетоалкилфосфиновых кислот типа 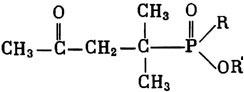 не оказывали антихолинэстеразного действия. В то же время они проявляли выраженные антидотные свойства но отношению к токсичным ФОС, угнетающим холинэстеразу. Ученые установили, в частности, что с помощью препаратов, у которых R=OCH3, a R'=CH3, можно спасти от гибели 80–100% подопытных животных, отравленных абсолютно смертельными дозами таких сильнодействующих ФОС, как октаметил и фосфакол. Что касается молекулярного механизма описанного эффекта «ФОС против ФОС», то каких-либо убедительных данных на этот счет пока не имеется. Известно только, что по отношению к холинэстеразе кетоалкилфосфонаты не проявляют заметного реактивирующего и защитного действия, но несколько повышают ее активность в организме. Представления о взаимосвязи нервного импульса, токсичных веществ и противоядий были бы неполными, если бы мы не рассмотрели структуру и функцию адренореактивных систем и токсикологические особенности соответствующих ядов и антидотов. Адренергические медиаторы и рецепторные структуры До сих пор речь шла о веществах, которые прямо или косвенно связаны с функцией одного из химических передатчиков нервного возбуждения — ацетилхолина. Иной молекулярный механизм лежит в основе действия ядов и антидотов, вмешивающихся в обмен адреналина и норадреналина, — медиаторов второго типа, получивших общее название катехоламинов. Биохимическим предшественником этих веществ является жизненно важная аминокислота тирозин: 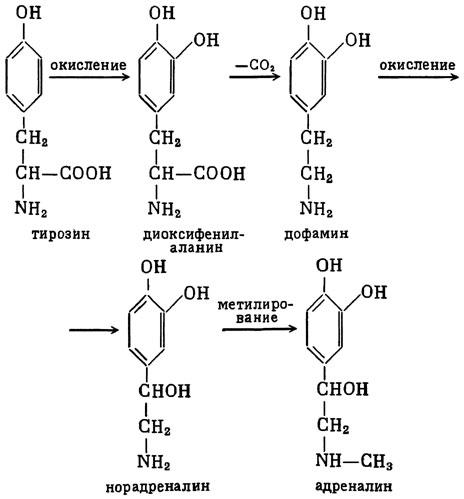 В настоящее время как медиатор рассматривается также дофамин, который, как видно из приведенной схемы, является промежуточным продуктом биосинтеза норадреналина и адреналина. Физиологическая роль этих передатчиков нервного импульса проявляется в центральных и периферических структурах, которые принято называть адренергическими в отличие от описанных ранее холинергических, связанных с ацетилхолином. Установлено, что катехоламины обладают выраженными свойствами регуляторов ряда процессов жизнедеятельности. Поэтому они, прежде всего адреналин, нередко рассматриваются как гормональные вещества. 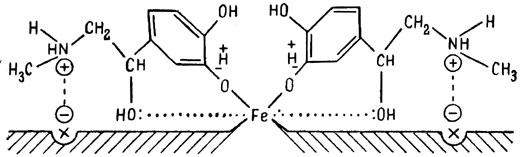 Рис. 9. Предполагаемая схема фиксации адреналина в активных центрах адренорецептивных структур (Комиссаров, 1959) Рис. 9. Предполагаемая схема фиксации адреналина в активных центрах адренорецептивных структур (Комиссаров, 1959) Не касаясь многих сложных и до конца еще не изученных сторон биохимической организации адренергических систем, можно полагать, что выброс норадреналина (точнее, его смеси с адреналином, именуемой симпатином) в синаптическую щель и последующее его взаимодействие с рецепторньми структурами (адренорецепторами) являются непременным условием нормального функционирования соответствующих отделов нервной системы. Адренорецепторы, с которыми взаимодействуют катехоламины, неоднородны. Это доказывается тем, что характер ответных реакций разных органов даже на один и тот же катехоламин может быть неодинаковым, а один и тот же орган может разнонаправленно реагировать на введение различных катехоламинов. В связи с этим по предложению американского ученого Алквиста (1948 г.) принято рассматривать 2 типа адренорецепторов: ?- и ?-адренорецепторы. Возбуждение ?-адренорецепторов связывается с сужением кровеносных сосудов, увеличением тонуса гладких мышц и усилением функции большинства соответствующих органов и структур, а стимуляция ?-адренорецепторов, наоборот, угнетает деятельность этих органов, расширяет кровеносные сосуды, повышает частоту и силу сокращений сердца. 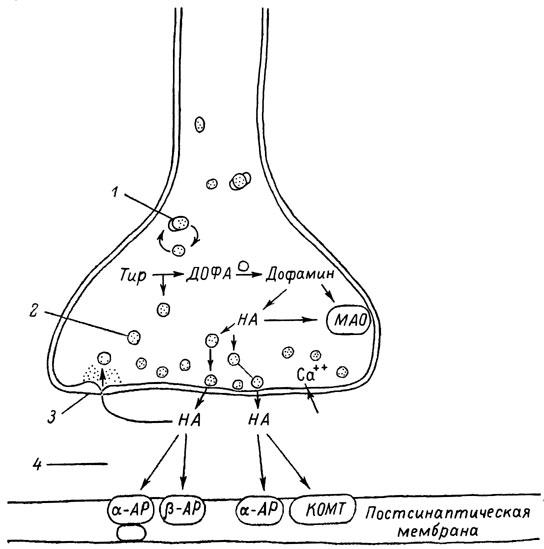 Рис. 10. Основные структурные элементы и схема функционирования адренер гического синапса (Голяков, Фишзон-Рысс, 1978). 1 — гранулы депонирования норадреналина (НА); 2 — синаптические пузырьки; з — обратный захват НА; 4 — сипаптическая щель; КОМТ — кате-хол-орто-метилтрансфераза; МАО — моноаминоксидаза; ДОФА — диокси-фенилалашш; Тир — тирозин; ?-АР и ?-AP — ?- и ?-адренорецепторы Рис. 10. Основные структурные элементы и схема функционирования адренер гического синапса (Голяков, Фишзон-Рысс, 1978). 1 — гранулы депонирования норадреналина (НА); 2 — синаптические пузырьки; з — обратный захват НА; 4 — сипаптическая щель; КОМТ — кате-хол-орто-метилтрансфераза; МАО — моноаминоксидаза; ДОФА — диокси-фенилалашш; Тир — тирозин; ?-АР и ?-AP — ?- и ?-адренорецепторы Согласно современным взглядам, адренорецепторы представляют собою металлопротеиды, имеющие двучленное строение с атомом железа или марганца в роли связующего звена, что позволяет одновременно фиксироваться на них 2 молекулам медиатора. По крайней мере такая структура с достаточным основанием приписывается ?-адренорецепторам (рис. 9). Мгновенная инактивация медиаторов в адренергических синапсах осуществляется большим числом ферментов,[88] из которых главное значение имеют моноаминоксидаза и катехоламин-орто-метилтрансфераза (КОМТ) (рис. 10). Данные ферменты катализируют окислительное дезаминирование катехоламинов. Биохимиками более изучена структура и функция моноаминоксидазы, которая в отличие от холинэстеразы является сложным металло-протеидом, имеющим в составе своей молекулы активную каталитическую часть особой химической структуры, именуемую в биохимии простетической, или коферментной и включающую пиридоксальфосфатную группировку и атомы меди. В свою очередь пиридоксальфосфат состоит из пиридоксина (витамина В6) и фосфорной кислоты. Согласно принятым данным,[89] именно пиридоксин и медь являются важнейшими компонентами активных каталитических центров моноаминоксидазы и ряда других ферментов, получивших общее название пиридоксалевых. Серотонин Результаты экспериментальных и клинических исследований позволяют теперь с достаточной определенностью говорить о существовании такого медиатора нервного возбуждения, биохимическая роль которого связана в основном с высшей нервной деятельностью. Речь идет о серотонине (5-окситриптамине), который считается химическим передатчиком нервных импульсов в центральных синапсах главным образом стволовой части головного мозга. Источником образования серотонина в организме является одна из жизненно важных аминокислот — триптофан, который под действием специфического фермента (оксидазы) превращается в 5-окситриптофан, а последний декарбоксилируется (теряет СО2) и превращается в серотонин: 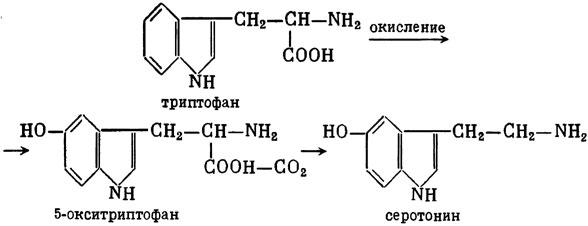 Понятно, что после выполнения медиаторной функции (т. е. после воздействия на соответствующие рецепторы) молекулы серотонина, подобно другим медиаторам, мгновенно инактивируются. Это происходит вследствие дезаминирования и окисления под влиянием фермента моноаминоксидазы. Следовательно, катехоламины и серотонин связывает общность путей биотрансформации. В этой связи небезынтересно сопоставить химическую структуру адреналина и серотонина. Если допустить, что этиламиновая группа адреналина изогнута, то по своей конфигурации его молекула представляет собою разорванный индольный гетероцикл:[90] 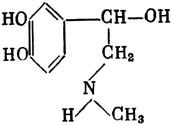 Основываясь на такой точке зрения, можно предположить, что биоструктуры, с которыми взаимодействует серотонин, близки по своему строению с адренорецепторами. Подобие химического строения серотонина с веществами, близкими к адреналину, имеет значение для понимания молекулярного механизма действия некоторых психотомиметических ядов. Гамма-аминомасляная кислота В последние 25 лет все большее внимание биохимиков, фармакологов, токсикологов привлекает еще один медиатор передачи импульсов в нервной системе — гамма-аминомасляная кислота (ГАМК). Этот медиатор является нормальным продуктом обмена веществ у млекопитающих и образуется из глутаминовой кислоты при действии пиридоксалевого фермента глутаматдекарбоксилазы: 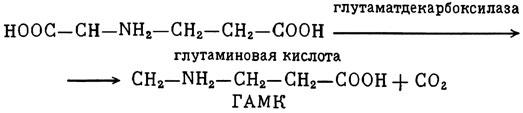 Теперь считается доказанным,[91] что ГАМК тормозит развитие и проведение импульсов в центральной нервной системе. Следовательно, можно полагать, что по своему биохимическому действию ГАМК есть антагонист тех эндогенных и экзогенных веществ, которые вызывают или стимулируют в нервной системе возбудительный процесс. Освобождающаяся при раздражении тормозных нервных структур (проводящих путей, нервов, клеточных скоплений) ГАМК преодолевает синаптическую щель и вступает во взаимодействие с рецепторами постсинаптической мембраны (ГАМК-рецепторами). По И. А. Сытинскому, ГАМК-рецептор — субклеточная структура (фосфолипидно-белковьй комплекс) постсинаптической мембраны с активными участками, облегчающими сорбцию медиатора на его поверхности. Не исключается, что ГАМК-рецепторы, подобно адренорецепторам, имеют двучленное строение и, следовательно, могут активироваться 2 молекулами медиатора. Медкаторное действие ГАМК в центральной нервной системе можно также объяснить сходством ее химического и пространственного строения с ацетилхолином: 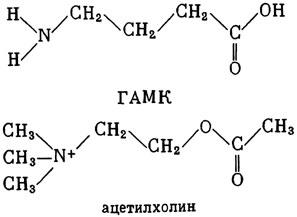 Поэтому учитывается возможность конкурентного антагонизма этих медиаторов в их действии на холинергические рецепторные структуры: взаимодействие с ГАМК защищает холинорецептор от стимулирующего влияния ацетилхолина. Как и другие медиаторы, ГАМК разрушается в постсинаптических структурах с помощью специфического катализатора. Им является фермент ГАМК-трансаминаза. Важно при этом иметь в виду, что при инактивации ГАМК вновь образуется ее предшественник — глутаминовая кислота. С другой стороны, закономерно, что блокада трансаминазы приводит к избытку ГАМК в синапсах. Яды — блокаторы пиридоксалевых ферментов Ряд ядов избирательно влияет на медиаторную функцию катехоламинов, серотонина и ГАМК. Один из них — сероуглерод (CS2) — высокотоксичное производное дитиокарбаминовой кислоты ( При попадании сероуглерода в организм через органы дыхания, кожу, зараженную пищу и воду возникают характерные явления нейроинтоксикации. Вначале отмечается возбуждение, нарушение координации движений, рвота, бред, а затем апатия, заторможенность, потеря памяти. В особо тяжелых случаях утрачивается сознание, угнетаются или даже исчезают зрачковые и сухожильные рефлексы, расстраивается дыхание и работа сердца. Работами ряда исследователей прежде всего советских (З. X. Черный, Ж. И. Абрамова и др.), с достоверностью установлено, что сероуглерод является специфическим ингибитором моноаминоксидазы. Это в свою очередь приводит к нарушению обмена биогенных аминов, в особенности окисления серотонина, накоплению его и других медиаторов в синапсах и к избыточной функции адрено-рецепторных структур. В свете такого механизма становится понятным найденный упомянутыми авторами при сероуглеродной интоксикации дефицит в организме витамина В6 и меди.[92] Надо иметь также в виду, что в организме CS2 связывается с другими биоструктурами, содержащими свободные сульфгидрильные и аминные группы. Образующимся при этом тиокарбаматным соединениям приписывается способность связывать биоэлементы, в том числе медь, и тем вторично нарушать функцию металлсодержащих ферментов (моноаминоксидазы и др.). Рассмотрим далее токсикологические особенности гидразина (N2H4) и его соединений, которые широко используются в промышленности (производство пластических масс, синтетических смол, каучука, красителей, взрывчатых веществ и др.), как ядохимикаты и химические реактивы, а также в клинической медицине при лечении туберкулеза, опухолевых заболеваний и в качестве психофармакологических средств. Кроме того, соединения гидразина как сильные восстановители применяются для стабилизации жиров, фруктовых соков и других продуктов, о некоторые из них используются как регуляторы роста растений. Несмотря на различия в химическом строении отдельных представителей этого класса соединений, их объединяют важнейшие признаки резорбтивного действия, которые мало зависят от путей поступления веществ в организм. При воздействии на организм больших доз гидразина и высокотоксичных веществ, синтезированных на его основе, на первый план выступают расстройства со стороны нервной системы: головная боль, возбуждение, судороги, потеря сознания, параличи, а также симптомы поражения печени. Теперь не вызывает сомнения, что основным первичным объектом их токсического воздействия на молекулярном уровне является уже знакомый нам фермент глутаматдекарбоксилаза, а во взаимодействие с ядами вступает ее кофермент — пиридоксальфосфат. Один из возможных механизмов такой реакции представлен в виде следующей схемы: 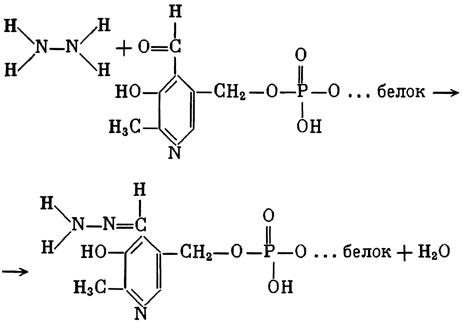 Таким образом, гидразин и его соединения блокируют реакцию превращения глутаминовой кислоты в ГАМК. Имеются данные, согласно которым ряд гидразинов связывает также ГАМК-трансаминазу и, кроме того, тормозит синтез пиридоксальфосфата. Тем самым еще больше усложняется цепь нарушений передачи импульсов в центральной нервной системе, главным образом — тормозных. Этим, однако, не ограничивается механизм влияния рассматриваемых ядов на организм. Считается доказанным, что многие из них, подобно сероуглероду, ингибируют и моноаминоксидазу, а следовательно, тормозят окислительное дезаминирование катехоламинов и серотонина и приводят к их накоплению в адренергических структурах. Это в свою очередь вызывает избыточную функцию адренорецепторов центральной нервной системы и, по-видимому, лежит в основе возникновения таких симптомов, как психомоторное возбуждение, эйфория и т. п. Итак, изложенное показывает, что многие ядовитые вещества вмешиваются в обмен биогенных аминов посредством блокирования пиридоксалевых ферментов. Основываясь на приведенных молекулярных механизмах и по аналогии с антихолинэстеразными ядами (см. с. 65), можно наметить несколько рациональных направлений антидотного воздействия на течение интоксикаций этими веществами. Однако существующие знания о структуре и биохимических свойствах адренореактивных систем и механизмах ингибирования пиридоксалевых ферментов позволяют реализовать (или предсказать реализацию в скором будущем) только те из них, при которых осуществляется: а) заместительное действие противоядия и б) влияние па функцию адренорецептивных структур Витамин В6, глутаминовая кислота и препараты меди как антидоты Витамины B6, или пиридоксин Витамин В6 (в виде хлорида пиридоксина) при острых отравлениях рекомендуется назначать в больших дозах.[95] В то же время при хронических формах отравлений, а также с профилактической целью применяются сравнительно небольшие его количества. Так, специально разработанная сотрудниками Ленинградского НИИ гигиены труда и профессиональных заболеваний инструкция[96] рекомендует для нормализации функциональных сдвигов в организме при хронической интоксикации сероуглеродом ежедневную дозу пиридоксина, равную 50 мг (1 мл 5%-ного раствора). С этой же целью инструкция предписывает комбинировать пиридоксин с глутаминовой кислотой. Механизм ее действия, как считают,[97] состоит прежде всего в химическом связывании яда и ускорении его выведения из организма. С другой стороны, в качестве антидота глутаминовая кислота должна рассматриваться и как биохимический предшественник ГАМК. Это ее свойство может оказаться полезным для специфического воздействия на течение и других интоксикаций, при которых нарушается обмен данного медиатора.[98] В последнее время для лечения различных нарушений функционирования центральной нервной системы все больше используется гаммалон — препарат гамма-амино-масляной кислоты. Его назначают внутрь в таблетках по 0,25 г и для введения в вену в виде 5%-ного раствора. Можно предположить, что этот препарат найдет практическое применение и как заместительное антидотное средство в первую очередь при гидразиновых отравлениях. Тот факт, что яды-ингибиторы моноаминоксидазы имеют химическое сродство к меди, входящей в состав данного фермента, давно наводил исследователей на мысль о применении этого микроэлемента при соответствующих отравлениях. И действительно, оказалось, что соединения меди, в частности уксусномедная соль, препятствуют развитию нарушений обмена веществ при сероуглеродной и гидразиновых интоксикациях, резко понижают накопление этих ядов и продуктов их превращений в организме, а также нормализуют метаболизм биогенных аминов. Все это, несомненно, свидетельствует о том, что препараты меди проявляют свойства антидотов. Вполне оправданы поэтому рекомендации по их практическому применению. Так, в настоящее время предписывается дополнительно вводить ацетат меди (совместно с пиридоксином) в рацион лечебно-профилактического питания лиц, контактирующих на производстве с сероуглеродом и другими дитиокарбаматами, а также обязательно включать его в комплекс антидотных средств при соответствующих интоксикациях. Адреноблокаторы Теоретически антидотньм эффектом при интоксикации ядами-ингибиторами пиридоксалевых ферментов должны обладать (по аналогии с холинолитиками) препараты, которые обратимо, но быстро и на достаточно длительный срок выключают адренорецепторы из механизма нервной передачи. Такими свойствами обладает большое число веществ, известных в фармакологии под названием адреноблокаторов. В зависимости от того, какой тип рецепторов преимущественно блокируется, их подразделяют на ?- и ?-адреноблокаторы. К первым относятся производные спорыньи (эрготамин, редергам), фентоламин, дибенамин, фенитрон. В числе вторых — пронеталол, индерал, дихлоризопротеренол, аптин, а также отечественный аналог индерала — анаприлин. Эти вещества, в особенности ?-адреноблокаторы, все более широко используются в клинике внутренних болезнен при различных патологических состояниях (например, при болезнях сердечно-сосудистой системы). Можно с уверенностью предсказать их использование в качестве антидотов для снятия явлений перевозбуждения адренорецепторов.[99] Не исключено, что адреноблокаторы окажутся полезными и для профилактики отравлений ингибиторами моноаминоксидазы, а также веществами, непосредственно стимулирующими функцию адренорецепторов. В предполагаемом механизме действия адреноблокирующих антидотов можно усмотреть и структурный компонент. Это, например, иллюстрируется сопоставлением строения молекулы адреналина и ?-адреноблокатора дихлоризопротеренола: 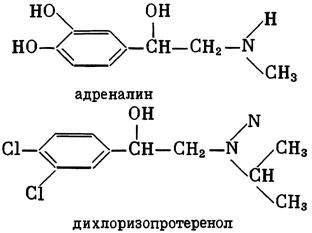 Естественно поэтому предположить, что конкурентный антагонизм этих веществ основывается на присоединении их молекул к одним и тем же звеньям адренорецепторной структуры. Таким образом, в перспективном плане адреноблокирующие соединения можно рассматривать как противоядия. Расширяющийся диапазон их терапевтического применения несомненно будет способствовать внедрению этих веществ в токсикологическую практику. Диэтиламид лизергиновой кислоты и его антагонисты В связи с разбирающимися токсическими нарушениями функционирования адренергических систем заслуживают отдельного рассмотрения некоторые психотомиметические вещества. Прежде всего это достаточно хорошо известный диэтиламид лизергиновой кислоты (ДЛК): 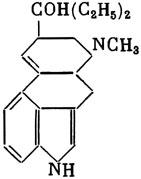 ДЛК — полусинтетическое вещество, полученное из ржаной спорыньи, которое завоевало за последние десятилетия в семействе психоядов главенствующее положение. Прошло уже более 35 лет с тех пор, как швейцарский фармаколог Гоффманн стал объектом случайного токсического воздействия ДЛК, несколько миллиграммов которого ему до этого удалось получить в виде кристаллического вещества. Поставив на себе опыт, Гоффман обнаружил, что в дозе 0,25 мг ДЛК вызывает многообразные остро протекающие расстройства психики: нарушается восприятие окружающей среды и своего тела, появляются различные галлюцинации, искажается эмоциональная деятельность и ориентация во времени и месте. В дальнейшем было выяснено, что ДЛК может привести также к развитию маниакального состояния, психомоторному возбуждению, бреду. В настоящее время считается, что действующая доза ДЛК для человека равна 0,07–0,15 мг, при этом психотоксический эффект отмечается в течение 4–6 ч.[100] На механизм действия психотомиметических ядов типа ДЛК на центральную нервную систему не существует однозначного взгляда. Считается, что в минимальных дозах эти вещества прекращают действие серотонина, вытесняя его с поверхности рецептора и вступая с ним, таким образом, в конкурентные отношения. С другой стороны, имеются данные за то, что большие дозы препаратов типа ДЛК, блокируя моноаминоксидазу, препятствуют нормальному окислению серотонина и катехоламинов и способствуют их накоплению в синапсах. Высказывается также мнение, что ДЛК повышает чувствительность центральных адренергических рецепторов к действию свободных медиаторов. Поэтому предполагается, что в зависимости от дозы ДЛК либо тормозится, либо усиливается действие медиаторов на адренорецепторы. Все это ведет к па рушению нормального проведения импульсов в адренергических структурах центральной нервной системы, что и рассматривается как биохимическая основа тех или иных психических расстройств.[101] В последние годы все чаще подчеркивается значение нарушений при интоксикации ДЛК нормального процесса биотрансформации адреналина и норадреналина. Это приводит к превращению их в адренохром и адренолютин — вещества, одновременно близкие по структуре к серотонину и к некоторым психотомиметическим агентам (псилоцину, буфотенину и другим производным аминокислоты триптамина): 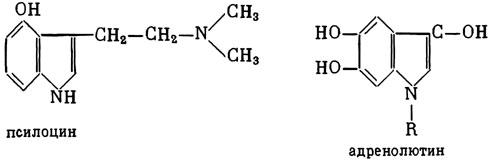 В эксперименте на людях было доказано, что адренохром и адренолютин вызывают психические расстройства. В этой связи интересно предположение о механизме действия другого психотомиметика — мескалина, алкалоида мексиканского кактуса лофофора. Оказалось, что в процессе превращений в организме норадреналина, в частности при его метилировании, могут образовываться вещества, весьма сходные по строению с мескалином. Вполне логично предположить, что и реакции метаболизма мескалина (тот же процесс метилирования) имеют связь со структурными превращениями данного медиатора и должны рассматриваться как звенья единого биохимического механизма нарушений психической деятельности. Изысканию антигаллюциногенных средств посвящено большое число как отечественных, так и зарубежных исследований. Так, весьма результативными оказались испытания различных фармакологических препаратов на крысах, у которых вызывался лизергиновый психоз.[102] В частности, было установлено, что предварительное (за 2 ч до отравления ДЛК) введение животным ?- и ?-адреноблокаторов заметно снижало у них выраженность нарушений поведения и двигательных расстройств, причем ?-адреноблокатор феноксибензамин оказался наиболее эффективным антагонистом ДЛК. ?-Адреноблокаторы пронеталол и индерал также нормализовали извращенное поведение животных, но в отличие от феноксибензамина почти не уменьшали двигательного возбуждения. В этой связи представляет интерес работа, выполненная О. Н. Давыдовой[103] в лаборатории профессора А. Н. Кудрина. В опытах на кошках была показана высокая противолизергиновая активность ?-адреноблокатора фенитрона. Действие этого препарата особенно было выражено при предварительном или одновременном его введении животным с токсичными дозами ДЛК. Автором было также установлено, что фенитрон является активным антагонистом другого весьма распространенного психотоксического вещества — гашиша. Из веществ, испытанных в качестве антагонистов ДЛК, следует назвать ?-метилпаратирозин, который оказался сильным ангибитором синтеза катехоламинов в головном мозге. Этот препарат отчетливо препятствовал развитию лизергиновой интоксикации при введении животным за несколько часов до отравления, а также нормализовал расстройства поведения, если его вводили животным через 20–30 мин после ДЛК. Пока еще нет опыта, позволяющего оценить практическую значимость представленных подходов к специфическому лекарственному воздействию на лизергиновые интоксикации. Однако определенные положительные сдвиги в этом направлении уже намечаются. Так, один из испытанных Диксоном (США) препаратов — дезерил, вмешивающийся в функцию центральных серотониновых структур, был с успехом использован для нормализации психической деятельности у больных.[104] Другой препарат подобного типа действия — мексамин (5-метокситриптамин):  Будучи структурным аналогом серотонина, мексамин в отличие от него преодолевает гематоэнцефалический барьер и проникает в головной мозг. В опытах на собаках профессор Г. И. Мильштейн[105] установил, что мексамин является эффективным антагонистом ДЛК. Это, по мнению автора, косвенно доказывает, что лизергиновый психоз является результатом нарушений в обмене серотонина. В этой связи практическую ценность имели данные Р. Б. Стрелкова и В. А. Хасабовой,[106] которые доказали (авторы ставили опыты на себе и на добровольцах) переносимость больших доз мексамина (до 6,5 мг/кг при приеме внутрь), что дало основания для дальнейшего испытания этого препарата как возможного антидота ДЛК. Пример с мексамином еще раз подтверждает научное значение известного положения, выдвинутого академиком АМН СССР В. М. Карасином: «Несомненно, что любая фармакологическая реакция имеет биохимический прототип: чуждый организму химический агент лишь в том случае может вступить в реакцию с той или иной биохимической структурой, если по своему химическому строению или поведению он сходен с каким-либо веществом, свойственным организму».[107] Примечания:1 Саноцкий И. В. Предупреждение вредных химических воздействий на человека — комплексная задача медицины, экологии, химии и техники. — ЖВХО, 1974, № 2, с. 125–142. 6 Подробно эта проблема рассмотрена в монографии прогрессивного американского публициста С. Херша «Химическое и биологическое оружие. Тайный арсенал Америки» (пер. с англ. М.: Воениздат, 1970). Работы но созданию высокотоксичных отравляюших веществ освещены также в книге Дж. Ротшильда «Оружие завтрашнего дня» (пер. с англ. М., Воениздат, 1966). 7 От токсикон — яд, логос — учение (греч.). В последние 20–25 лет во всем мире, в том числе и в СССР, токсикология достигла значительного развития. С учетом важного значения этой науки для охраны здоровья населения и развития народного хозяйства в 1981 г. было создано Всесоюзное научное общество токсикологов (ранее токсикологи, фармакологи и биохимики были объединены в одном обществе). 8 Пирузян Л. А., Маленков А. Г., Баренбойм Г. М. Химические аспекты деятельности человечества и охрана природы. — Природа, 1980, № 3, с. 11. 9 Рудаки А. Касыды. — В кн.: Ирано-таджикская поэзия/ Пер. с фарси. М.: Худож. лит., 1974, с. 23. (Сер. Б-ка всемир. лит.). 10 (Лужников Е. А., Дагаее В. Н., Фарсов Н. Н. Основы реаниматологии при острых отравлениях. М.: Медицина, 1977. 69 Франке З. Химия отравляющих веществ/ Пер. с пом. под ред. И. Л. Кнунянца и Р. Н. Стерлина. М.: Химия, 1973, с. 248. 70 Ацетилкоэнзим А — соединенно уксусной кислоты с нуклеотпи дом, включающим несколько аминокислот и активную SН-группу. Отщепляя ацетат, идущий на построение молекулы ацотилхолина, он превращается в коэнзим А. 71 Глебов Р. Н., Примаковский Г. Н, Функциональная биохимия синапсов. М.: Медицина, 1978. 72 Согласно установившейся точке зрения, возникновение разности потенциалов между наружной и внутренней сторонами поверхностного сдоя клетки обусловлено неравномерным распределением ионов Na+ и К+ по обе стороны клеточной мембраны. При этом компенсирующий поток ионов К+, направленный в обратную сторону при воздействии медиатора на постсинантическую мембрану, несколько запаздывает, что приводит на короткое время к обеднению внешней поверхности клетки положительными ионами. 73 Розенгарт В. И. Холинэстеразы. Функциональная роль и клиническое значение. — В кн.: Проблемы медицинской химии. М.: Медицина, 1973, с. 66–104. 74 Прозоровский В. Б. Вопросы механизма действия и возрастной токсикологии антихолинэетеразных средств. Автореф. докт. дис. Л., 1969. 75 (Ганглиоблокаторы — вещества, избирательно тормозящие передачу нервных импульсов в вегетативных узлах (ганглияхвследствие блокады их холинорецепторов. 76 Каган Ю. С. Актуальные вопросы токсикологии фосфорорганических инсектицидов. — В кн.: Гигиена, токсикология и клиника новых инсектофунгицидов. М.: Медгиз, 1959, с. 187–217. 77 Кузнецов С. Г., Голиков С. И. Синтетические атропиноподобные вещества. Л.: Медгиз, 1962. Эти взгляды на взаимодействие холинолитика с холинорецептором в дальнейшем были развиты и дополнены (Голиков С. Н., Кузнецов С. Г. Современные представления о природе холинорецептора. — Вестн, АМН, 1970, № 2, с. 67–85). 78 Лебедева Д. П. О действий холпнолитиков группы атропина в комплексе с поливинолом. — Фармакол. и токсикол., 1971, № 6, с. 657–600. 79 Задолго до выделения атропина в чистом виде были известны случаи отравления людей и животных ягодами красавки. Например, с целью предупреждения интоксикаций еще в конце XVIII в. в Австрии было издано несколько циркуляров с подробным описанием этого растения и его ядовитых свойств. 80 Hauschild F. Pharmakologie und Grundlagen der Toxikologie. Leipzig, 1960. 81 Мильштейн Г. И., Спивая Л. И. Психотомиметики. Л.: Медицина, 1971. 82 Однако антидотное действие оксимов не обязательно связано с вовлечением в процесс реактивации анионного центра холинэстеразы, так как существует ряд веществ из класса оксимов, в молекулах которых не содержится четвертичный азот, но которые тем не менее проявляют выраженный реактивирующий эффект (изонитрозин, моноизонитрозоацетон и др.). 83 Koelle G. В. Cholinesterases and anticholinesterase agents. Handbuch der exper. Pharmakologie. Berlin, 1963, Bd 15. 84 Шеин Г. К. Сочетанное применение изонитрозина и ТМБ-4 при отравлении фосфорорганическими соединениями. — Фармакол, и токсикол., 1967, № 4, с. 491–493. 85 Голиков С. Н., Заугольников С. Д. Реактиваторы холинэстераз. Л.: Медицина, 1970. 86 Алексашина З. А. Токсикология, экспериментальная терапия и лекарственная профилактика отравлений Севаном. Автореф, канд. дис. Mинск, 1969. 87 Арбузов Б. А., Визель А. О., Ивановская К. М. Синтез и новые биологические эффекты ФОС с низкой токсичностью, — ДАН СССР, 1968, т. 182, с. 101–104. 88 Так, профессор А. М. Утевский указывает на наличие 6 групп ферментов, каждая из которых ответственна за биохимические превращения отдельных составных частей молекул катехоламинов (Утевский А. М. Обмен катехоламинов и его функциональное значение. — В кн.: Адреналин и норадреналин. М.: Наука, 1964, с. 8–18). 89 Горкин В. З. Моноаминооксидазы (современные представления о природе и физиологической роли). — В кн.: Биогенные амины. Тр. 1-го Моск, мед. ин-та/ Под ред. В, В. Меньшикова. М., 1967, т. 52, с, 146–161. 90 Салямон Л. С., Галлюциногены. — В кн.: Руководство по фармакологии. Л.: Медгиз, 1961, т. 1, с. 439–440. 91 Сытинский И. А., Гамма-аминомасляная кислота — медиатор торможения в нервной системе, — Природа, 1973, № 1, с. 20–29. 92 Есть еще немало распространенных в промышленности производных тио- и дитиокарбаминовой кислоты (карбатион, цинеб и др.), которые, как и сероуглерод, являются ингибиторами моно-аминоксидазы и вызывают во многом сходную с ним картину отравления. Подобным же образом действует на организм такой известный в химическом производстве продукт, как трихлор-этилен. 93 Черный 3. X. Некоторые вопросы патогенеза сероуглеродной интоксикации и подходы к ее специфической терапии. Автореф, канд. дис. Л., 1969. 94 Лазарев Н. В. Основные принципы лечения острых отравлений. Л., 1944, с. 108. 95 Например, при тяжелой гидразиновой интоксикации вводят одномоментно до 1.5 г этого препарата, причем такая доза может назначаться повторно (Back К. С., Pinkerton M. К., Thomas A. A. Therapy of acute UDMH intoxication. — Aerospace Med., 1963, v. 34, p. 1001–1004). 96 Инструкция по применению витамина В6 и глутаминовой кислоты для лечения хронических сероуглеродных интоксикаций. Л.: Ленинград. НИИ гигиены труда и проф. заболеваний, 1968. 97 Абрамова Ж. И. Вопросы специфической профилактики некоторых профессиональных интоксикаций. — В кн.: Матер. науч. сессии, посвящ. 40-летию НИИ гигиены труда и проф. заболеваний. Л., 1964, с. 102–105. 98 Глутаминовую кислоту рекомендуется применять внутрь в виде кальциевой соли по 20–50 мг или же внутривенно по 10 мл 10%-ного раствора. 99 Защитное действие ?-адреноблокаторов нередко сопровождается побочными эффектами, что связывается с необратимостью их реагирования с рецепторами ряда внутренних органов и центральной нервной системы. 100 Столь высокая токсичность ДЛК объясняет особый интерес, который проявляют к этому веществу военные круги стран НАТО. Так, в армии США производные лизергиновой кислоты рассматриваются в качестве отравляющих веществ, временно выводящих войска из строя. 101 Мильштейн Г. И., Спивак Л. И. Психотомиметики. Л.: Медицина, 1971. 102 Цит. по: Давыдова О. Н., Кудрин А. Н. Антагонисты диэтиламида лизергиновой кислоты. — Фармакол. и токсикол., 1971, № 5, с. 629–631. 103 Давыдова О. Н. Влияние гашиша и ДЛК на поведение экспериментальных животных и изыскание антагонистов этих веществ. Автореф. канд. дис. М., 1968. 104 Камбарова Д. К. Центральные биохимические системы и патология. — В кн.: Проблемы физиологии и патологии высшей нервной деятельности. Л., 1970, вып. 4, с. 185. 105 Мильштейн Г. И. Влияние мексамина на центральные эффекты диэтиламида лизергиновой кислоты. — Фармакол. и токсикол., 1966, № 6, с. 662–665. 106 Стрелков Р. В., Хасабова В. А. К токсикологии мексамина. — Фармакол. и токсикол., 1968, № 6, с. 731–733. 107 Карасик В. М. Аллостерические эффекты в фармакологических реакциях холинергических структур. — В кн.: X съезд Всесоюз. физиол. о-ва им. И. П. Павлова. Ереван: Наука, 1964, т. 1, с. 64. |
|
|||
|
Главная | Контакты | Прислать материал | Добавить в избранное | Сообщить об ошибке |
||||
|
|
||||
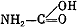 ) карбаматов. Оказалось, что реактиваторы из класса оксимов почти не оказывают влиякая на течение интоксикации севином, и это связывается с тем, что данный яд блокирует холинэстеразу не посредством фосфорилирования, а за счет взаимодействия с эстеразным пунктом фермента его карбаминовой группировки (так называемое карбаминирование). В то же время холинолитические препараты (атропин, тропацин и др.) проявляют при отравлении карбаматами выраженное антидотное действие, что подчеркивает ведущее значение избыточной функции холинорецепторов при интоксикациях антихолинэстеразньми веществами разного химического строения.
) карбаматов. Оказалось, что реактиваторы из класса оксимов почти не оказывают влиякая на течение интоксикации севином, и это связывается с тем, что данный яд блокирует холинэстеразу не посредством фосфорилирования, а за счет взаимодействия с эстеразным пунктом фермента его карбаминовой группировки (так называемое карбаминирование). В то же время холинолитические препараты (атропин, тропацин и др.) проявляют при отравлении карбаматами выраженное антидотное действие, что подчеркивает ведущее значение избыточной функции холинорецепторов при интоксикациях антихолинэстеразньми веществами разного химического строения.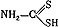 ), широко применяющееся в производстве вискозных волокон, целлофана, ядохимикатов, четыреххлористого углерода, а также в качестве растворителя.
), широко применяющееся в производстве вискозных волокон, целлофана, ядохимикатов, четыреххлористого углерода, а также в качестве растворителя.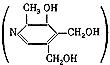 , согласно экспериментальным и клиническим данным, проявляет выраженные антидотные свойства при отравлениях сероуглеродом, другими дитиокарбаматами и трихлорэтиленом. Например, при сероуглеродной интоксикации под воздействием пиридоксина наблюдается снижение циркуляции яда в крови и повышение его выделения с мочой, что, по-видимому, говорит об образовании нетоксичного комплекса сероуглерода с антидотом.
, согласно экспериментальным и клиническим данным, проявляет выраженные антидотные свойства при отравлениях сероуглеродом, другими дитиокарбаматами и трихлорэтиленом. Например, при сероуглеродной интоксикации под воздействием пиридоксина наблюдается снижение циркуляции яда в крови и повышение его выделения с мочой, что, по-видимому, говорит об образовании нетоксичного комплекса сероуглерода с антидотом.