|
||||
|
Глава 2 Земная жизнь Судьбы человечества неотделимы от истории земной биосферы. Чтобы оценить свое место в мире, нам необходимо правильно понимать историю живого, однако в ней немало неясных и спорных страниц. Успехи эволюционной биологии и сопряженных с ней наук в прошлом веке заполнили множество «белых пятен» в летописи земной жизни, однако дискуссии по ее узловым моментам все еще далеки от завершения. Мы рассмотрим здесь: 1) современные гипотезы относительно возникновения органической жизни и ее возможной связи с пребыванием Солнечной системы поблизости от центра Вселенной в эпоху биогенеза; 2) общие сведения о глобальной тектонике плит, суперконтинентальном цикле, его влиянии на историю климата, экологию и направленность биологической эволюции; 3) современные сведения об узловых пунктах биологической эволюции, массовых вымираниях и расцветах новых форм организмов, а также о просматривающихся за этим повторяющихся, закономерных чертах эволюционного процесса, который выражается в чередовании мобильных и маломобильных фаун под влиянием оледенений и межледниковий; 4) новейшие данные о возникновении предков человека как о закономерном эпизоде биологической эволюции на Земле, в которой наши пращуры, гоминины, выступили единственными представителями очередной маломобильной фауны млекопитающих, вызванной к жизни одним из кайнозойских потеплений. 2.1. Биогенез Возраст нашей планеты достигает 4,55 млрд лет [805], а возраст Солнечной системы – 4,566 ± 0,002 млрд лет [395, с. 144]. В эпоху геогенеза (т. е. в период достижения Землей нынешней массы с металлическим ядром и примитивной атмосферой 4,51–4,45 млрд лет назад) расстояние Солнечной системы от центра Вселенной составляло 200,1 млн световых лет (при постоянной Хаббла в 57,4 км/с на мегапарсек и местной плотности Вселенной ?loc= 0,918), а в эпоху гелиогенеза (т. е. в период формирования Солнца) расстояние нашего новорожденного светила от центра Вселенной не превышало 198,8 млн светолет (при H0 = 57,5 км/с на мегапарсек и ?loc= 0,92). Новорожденная Земля пребывала тогда в относительно близких и древних окрестностях центра Вселенной. Это астрофизическое обстоятельство весьма существенно для биогенеза. Благодаря активному обмену веществом и энергией со средой живые системы устойчивы против Второго начала термодинамики. Они не только противодействуют росту энтропии внутри себя, но и генерируют порядок, т. е. создают негэнтропию. В этой связи встает вопрос: каким образом в расширяющейся Вселенной с растущей энтропией зародилась негэнтропийная жизнь? Проще говоря, живые существа не только умеют постоять за себя перед лицом множества стихий, но и воспроизводят себе подобных (генерируют порядок). Происхождение этих способностей предполагает некую «инкубацию» в тепличных условиях. Однако наша остывающая Вселенная никогда не обещала ничего комфортного в этом смысле. Земной биогенез пока непостижим [681]. Возможно, жизнь принесена на Землю с другой планеты посредством странствующего небесного тела (кометы, астероида, метеорита) [282; 306; 754]; а также [189; 230; 251; 301; 562; 594]; cp. [525]. Паракодоновый генетический код изученных организмов старше классического [300], что может означать, что исходный код сложился вне Земли, а на Земле переадаптировался. Такой вариант развития событий мыслим, что не снимает вопроса о происхождении жизни, пусть и не на Земле. На наш взгляд, не исключен следующий сценарий. На ранних стадиях эволюции Вселенной в ее центральных областях сохранялись реликты былой негэнтропии, оставшиеся от сингулярности, в то время как на периферии они рассеивались быстрее (параллельно падению постоянной Хаббла). Условия, благоприятные для зарождения жизни, отвечают среде горячих глубоководных источников с температурой 113° С и более [278; 342; 427; 446; 465; 484; 525; 594]; cp. [212; 339; 571; 614]. Подобная обстановка присуща подводным рифтам с высокими давлениями и кипящим химическим рассолом. Однако в наших океанах такие точки являются оазисами. Вероятность завоевания ими целой землеподобной планеты растет ближе к центру Вселенной, и вот почему. В окрестностях центра Вселенной велика средняя плотность вещества (см. приложение 7) и, соответственно, значительны гравитационные силы, что сообщает землеподобным планетам данного региона более высокие давления атмосферы и гидросферы. Подобные условия поднимают температуру кипения воды. Внушительное планетарное тяготение сильнее разогревает недра планеты, вызывает бурную мантийную конвекцию (тепловое перемешивание) и активную тектонику плит с обильными рифтами (см. разд. 2.2). На планете с такими условиями формируется плотный океан, густо испещренный глубоководными рифтами, где химически насыщенные воды кипят при температурах, перекрывающих земные, что повышает шансы пребиотического твердофазового синтеза нуклеотидов и аминокислот на минеральных поверхностях [321; 427] – как полагают, существовал вариант самопроизвольного возникновения материала нуклеиновых кислот (дезоксирибонуклеиновых – ДНК ядра клетки, и рибонуклеиновых – РНК цитоплазмы) и аминокислот (строительных блоков для белков) благодаря каталитической помощи некоторых минеральных поверхностей. Планета указанного типа имела бы предпочтительные возможности для биогенеза. Можно представить, как на заре Вселенной в звездных системах близ ядер галактик, превосходящих по возрасту шаровые скопления на 1–2 млрд лет [270, с. 25], или в самих шаровых скоплениях, поскольку ядра галактик с затравочной «черной дырой» в центре чересчур турбулентны, чтобы служить колыбелью жизни, – в какой-то планетной системе, парящей в центре Вселенной, на планете земной группы, отличающейся вышеописанными свойствами, зародилась органическая жизнь. Первоначально она могла существовать в виде «мира РНК» [353; 754], когда наследственные свойства организмов кодировались только рибонуклеиновой кислотой (РНК), а дезоксирибонуклеиновая кислота (ДНК) еще ожидала своего часа. В известном смысле подобный мир напоминал бы планету вирусов, обходящихся одной РНК: у нас они паразитируют в клетках более сложных существ – там питались бы химическими рассолами. Метеоритный материал с такой планеты мог бы перенести тамошние вирионы (закуклившиеся в белковую оболочку спящие вирусы) в Солнечную систему и основать «мир РНК» на Земле. Панспермия (космический посев жизни) мыслится и на стадии ДНК-содержащих бактерий. Попав к нам, живой инопланетный материал угодил бы в нищенскую среду, тем более что молодое Солнце в гадейской эре (4,5–3,8 млрд лет назад) давало лишь 70% нынешних света и тепла. В инфракрасной (тепловой) части спектра недостача восполнялась парниковым эффектом за счет аммония в гадее, а впоследствии за счет метана и особенно двуокиси углерода [208; 450; 601; 691]. Чужеродной теплолюбивой жизни пришлось бы укрыться в оазисах глубоководных горячих источников. Для живых существ это было бы равносильно переселению из более продуктивной (жаркой) в менее продуктивную (просто горячую) среду, что закалило гипотетических мигрантов и подготовило их к земной эволюции вполне определенной направленности, зависящей от геологической и климатической истории планеты. 2.2. Геология Сущность господствующей в геологии глобальной тектоники плит [65; 77; 96] заключается в следующем. Земля состоит из железо-никелевого, в основном железного ядра (16% объема и 1/3 массы планеты), которое окружено промежуточной оболочкой, мантией (около 2/3 массы планеты), а ее разуплотненный верхний слой, астеносфера, подстилает твердую земную кору, литосферу. Последняя разбита на огромные базальтовые блоки, литосферные плиты. В некоторые из этих плит впаяны обширные, менее плотные гранитные образования, представляющие собой поверхностные континенты. Дно океанов образовано более плотными базальтами. Разогреваясь от радиоактивного распада расщепляющихся элементов, содержащее их мантийное вещество разуплотняется и всплывает, приводя в движение астеносферу, остывает и вновь погружается в недра Земли (это называется мантийной конвекцией). Текущая астеносфера тянет с собой блоки базальтовой коры. В местах, где они расходятся, вещество астеносферы поступает наружу, формируя срединно-океанические хребты с рифтами (центральными впадинами) и наращивая отступающие базальтовые плиты (поэтому ближе к хребтам они всегда моложе). Происходит раздвижение (спрединг) океанического дна. Противоположные края плит вслед за конвективными течениями погружаются в астеносферу и тают в мантии – при этом плита ныряет под встречную (субдукция). В мантии тонут лишь тяжелые, базальтовые части плит, а гранитные (континентальные) из-за меньшего удельного веса остаются на поверхности планеты и даже наращиваются легкими изверженными породами в местах субдукции (островными дугами вроде Курильских островов). По своему происхождению все материки являются пакетами спрессованных островных дуг, наслоившихся друг на друга в течение 4,4 млрд лет плитовой тектоники на Земле. Континенты вместе с плитами медленно блуждают по поверхности планеты (континентальный дрейф). По ходу движения они сминаются в гигантские складки (горы вроде Анд). Сталкиваясь между собой, материки тоже сминаются, образуя горы (вроде Урала). Они могут спаиваться в более крупные континентальные образования, способные объединить даже все гранитные массивы в единый материк (Пангею, Мегагею и т. п.), который впоследствии раскалывается под действием мантийной конвекции и разъезжается осколками в разные стороны. По существующим представлениям [589; 590; 418], данный процесс носит циклический характер. Примерно раз в 500 млн лет материковые массивы, перемещающиеся на своих базальтовых плитах, сближаются над нисходящими токами мантийной конвекции, сталкиваются и спаиваются горными поясами в единый суперконтинент. Его окружает безбрежный океан, базальтовое дно которого все время обновляется вследствие субдукции и спрединга. Сверхматерик душит нисходящий ток конвекции под собой и существует около 100 млн лет. За этот интервал происходит перестройка конвекции в мантии, и под суперконтинентом начинает скапливаться тепло, образуя восходящий ток конвекции. Как следствие, над восходящим током конвекции в сверхматерике закладываются рифты (тепловые трещины), и спустя 40 млн лет (140 от начала цикла) единый континентальный массив раскалывается. Родившиеся из его осколков материки начинают дрейфовать в разные стороны. Между ними возникают новые, молодые океаны (вроде Атлантического) с новым океаническим дном базальтовой породы (220 млн лет от начала цикла). Спустя еще 100 млн лет (320 от начала цикла) континенты максимально удаляются друг от друга. Однако тяжелая базальтовая океаническая кора не может более 200 млн лет удерживаться на плаву над астеносферой (океанической коры старше 200 млн лет в современных океанах не известно, в том числе в «вечном» Тихом океане [204]). Максимальный возраст присущ базальтовой коре вдоль тыльных границ расходящихся материков. От старости она проваливается в мантию, тянет континенты на себя, и они приходят в попятное движение, поскольку конвекция уже перестроилась под влиянием обрушившейся океанической коры (420 млн лет от начала цикла). Материки начинают сближаться и в конце концов сталкиваются, образуя новый суперконтинент спустя 500 млн лет от начала цикла (цифры здесь ориентировочны). Затем все возобновляется, и часто на месте спаек материков (хотя и не всегда) зарождаются новые рифты, раскалывающие суперконтинент… Таким образом в суперконтинентальном цикле по 200 млн лет уходит на рождение и субдукцию дна молодых океанов, плюс около 100 млн лет на жизнь сверхматерика (что тоже ориентировочно). Около 250 млн лет назад образовался суперконтинент Пангея (потом он распался, так что современное человечество обитает на его осколках-материках). Однако ему предшествовали и другие сверхматерики, отмеченные 565 ± 5–549/543 млн лет назад (эдиакарская фауна экваториального суперконтинента [203; 258; 293; 381]), а также 1–1,1; 1,5–1,7; 2–2,1; 2,5–2,7 млрд лет назад – последняя дата отвечает рубежу протерозойского (2,5–0,544 млрд лет назад) и архейского (3,8–2,5 млрд лет назад) эонов. Отметим, что перемещающиеся по поверхности планеты континентальные породы существовали уже в раннем гадее 4,404 ± 0,008 млрд лет назад [805]; а также [204; 223; 395]; cp. [419]. Следовательно, в столь отдаленное время (всего спустя ок. 145 млн лет после образования Земли) имели место материковый дрейф и тектоника плит; ср. [459; 460; 478]. Старейшие орогены (горообразования) датированы св. 3,0; 2,8–2,6; 2,0–1,9; 1,90–1,88; 1,88–1,84; 1,78–1,72; 1,67–1,63 млрд лет назад [418; 459]. Часть этих орогенов связана с образованием сверхматериков, часть – с эпизодом расхождения континентов. Заметим, что суперконтинентальный цикл существенен для истории земного климата. В периоды суперконтинентального цикла Земля переживает по две пары эпизодов с крайними гидростатическими состояниями минимальных и максимальных уровней Мирового океана. В эпоху существования сверхматерика и в эпоху предельного расхождения континентов (как в настоящее время) на планете имеются наибольшие площади глубоководного океанического дна. В результате суша вздымается, уровень Мирового океана падает, а на Земле образуются максимальные по площади материковые поверхности. Последние же, в отличие от поверхности вод, обладают высоким альбедо (т. е. отражательной способностью) и мешают нагреванию планеты Солнцем. Если при этом суша попадает в околополярные области, на ней развиваются покровные оледенения [683; 70, с. 17–18]. Напротив, после раскола суперконтинента, когда начинается дрейф материков друг от друга, и после развития встречных движений континентов, когда они сближаются, океаны между ними мелки, уровень Мирового океана повышается, суша на окраинах материков затопляется, а планетарное альбедо снижается, позволяя Солнцу разогревать Землю. Наступает теплый и влажный климатический период – межледниковье. Иными словами, за время суперконтинентального цикла Земля закономерно переживает два оледенения и два межледниковья. Оледенения случались со средней периодичностью в 150 млн лет [42, с. 24, табл. 1; с. 30–35]. В эпоху оледенения на Земле устанавливается умеренный сухой климат, и такая обстановка не способствует высокой биопродуктивности земной экосреды. Растительность в среднем становится скудной и разреженной в пространстве. Напротив, в эпохи межледниковья климат становится теплым и влажным, что благоприятствует высокой биопродуктивности экосреды; растительность, как правило, богата и скучена в пространстве. Когда при оледенениях растительность скудна, разрежена, и вообще пищевые ресурсы бедны и рассеяны, животным приходится собирать корм с больших пространств. В такие периоды всем организмам эволюционно выгодно быть подвижными, высокомобильными, хотя это сопряжено с большими потерями энергии на передвижение. В противоположность этому, при межледниковьях растительность изобильна и скучена, как и все пищевые ресурсы, так что животным не надо искать пищу по обширным территориям. В такие эпохи животным и всем организмам вообще ни к чему быть высокомобильными, поскольку эволюционно не выгодно тратить энергию на лишние передвижения. Проще говоря, наиболее успешные живые организмы при оледенениях становятся высокоподвижными, а при межледниковьях – наоборот. Подобная закономерность представляется схематичной, однако ее можно проиллюстрировать. Допустим, на Земле стоит ледниковый период, в нем господствуют подвижные животные, а на вершине пищевой пирамиды царят высокомобильные хищники, которые, не считаясь с энергозатратами, без устали патрулируют свою экологическую нишу. Пища там скудна и ее необходимо потреблять без остановки, чтобы она не досталась конкурентам. При этом в тени господствующих высокомобильных существ обычно прозябают малоподвижные организмы, на которых никто не обращает внимания. Но вот оледенение заканчивается, становится тепло и влажно. Растения множатся и скучиваются в пространстве (как ныне свойственно им в тропиках и субтропиках). Наступает эпоха обильных кормовых угодий, и больше не требуется обшаривать значительные территории в поисках еды. Однако подвижные господа прежней, холодной эпохи в силу закона Долло о необратимости эволюции уже не в силах изменить свою физическую и нейрофизиологическую организацию и продолжают взад-вперед сновать по эконише, без толку тратя энергию на суету. Пока эти трудяги прочесывают один край биома, на другом краю «расправляют крылья» недавние аутсайдеры ледниковых времен – малоподвижные организмы. Они не торопясь и экономя энергию потребляют обильную пищу, так что, когда из рейда по экосистеме возвращаются высокомобильные существа, им питаться уже нечем. Пока они удивляются превратностям судьбы, ситуация повторяется в других уголках экосреды. Господа эпохи оледенения приходят в упадок, а воцаряются их малоподвижные конкуренты. Когда оледенение возвращается вновь, картина воспроизводится с противоположным знаком. Пища вновь скудна и редка. Ее недостает малоподвижным господам межледниковья, а осваивать большие пространства они не в силах. Уцелевшие остатки высокоподвижной фауны, коротающие теплые времена на задворках экосреды (как млекопитающие мезозоя; см. разд. 2.3), поднимают голову, обнаруживают, что снова стали актуальны, и приступают к подвижному патрулированию биоты. Недавние малоподвижные господа теплолюбивой жизни частью не выдерживают нового направления конкуренции и вымирают, а частью уходят в тень – до будущих теплых времен. Многочисленные палеонтологические примеры сказанному мы приводим ниже (см. разд. 2.3). Таким образом, можно увидеть определенную тенденцию земной эволюции, которая направляется экологией, та, в свою очередь, климатом, а он – дрейфом материков, т. е. циклическим геологическим фактором. Продемонстрируем на фактах, что подобная эволюционная тенденция действительно реализовалась в истории земной жизни. 2.3. Эволюция Археозой (4,55-2,5 млрд лет назад) Согласно концепции абиогенеза (о химической эволюции жизни из неорганического материала), процесс начался в глубоководных гипертермальных источниках 4,2–4,0 млрд лет назад, а 4,0–3,7 млрд лет назад достиг поверхностных вод [525]. Жидкая вода возникла на Земле 4,3 млрд лет назад [575]. Следы ископаемых организмов появляются в палеонтологической летописи в позднем гадее св. 3,85 млрд лет назад [422], а также [156; 225; 282; 465; 574; 580; 600; 677; 678; 788]. Отметим, что по молекулярно-генетическим данным возраст первых организмов оценивается приблизительно в 1,8 млрд лет [578], что вдвое уступает палеонтологическому возрасту; по-видимому, 1,8 млрд лет назад – это возраст эволюционного расхождения тех организмов, чьи прямые потомки дожили до наших дней, поскольку молекулярно-генетический возраст, например, нескольких родственных видов живых существ определяется до момента разветвления эволюционных путей их предков. Под влиянием мобилизующих колебаний продуктивности среды древние микроорганизмы развили пищевую автономность (автотрофность), чтобы не зависеть от внешних ресурсов в неурожайные (ледниковые) периоды. Около 3,8–3,5 (или 3,5; 3,465; 3,416) млрд лет назад цианобактерии освоили фотосинтез, т. е. способность синтезировать углеводы, аминокислоты, белки, пигменты и другие соединения под действием солнечного света [232; 788; 677; 678; 753]; а также [399; 465; 600]. Побочным продуктом фотосинтеза является свободный кислород. Однако поначалу он практически весь уходил на окисление железа, так что 2,8 млрд лет назад свободного кислорода в атмосфере почти не было [465, с. 2112]. Лишь 2,32 млрд лет назад наметился его атмосферный рост [181]. Чтобы добиться еще большей независимости от пищевых ресурсов среды, микроорганизмы путем симбиотического включения в свои тельца других одноклеточных существ обзавелись св. 2,8 млрд лет назад митохондриями [470], служащими внутренними источниками энергии для клеток, что подобно аккумуляторам повышало их мобильность. Митохондрии поначалу были самостоятельными микроорганизмами, а впоследствии поселились в тельцах других микроорганизмов и стали служить им «энергетическими подстанциями». Примерно тогда же, св. 2,7 млрд лет назад, прокариоты (организмы без клеточного ядра) путем аналогичного симбиоза поселили в своих тельцах другие микроорганизмы, которые стали играть для них роль клеточных ядер. В результате прокариоты стали эукариотами, организмами с клеточными ядрами [213; 294; 322; 465; 718]. Этим способом содержащийся в клеточном ядре геном приобрел как бы скафандр в виде материнской клетки, что повысило его независимость от условий среды и ее перепадов, а следовательно подняло его мобильность. Поэтому хронологическая близость возникновения митохондрий и клеточных ядер может отвечать климатическому пессимуму, возможно связанному с оледенением, ознаменовавшим расхождение материков 2,85 млрд лет назад. Отметим, что в силу той же логики появление белковой оболочки (вириона) у РНК-вирусов тоже могло быть откликом на климатический или другой пессимум в их истории, однако палеонтология вирусов не развита. Протерозой (2,5-0,544 млрд лет назад) После распада позднеархейского суперконтинента Мегагеи, образовавшегося и расколовшегося 2,7–2,5 млрд лет назад, имели место три Гуронских ледниковых периода (Рамсей-Лейк, Брюс и Гоуганда) в интервале 2,5–2,2 млрд лет назад. Они, вероятно, связаны с эпизодом сильного расхождения материков (см. разд. 2.2). Падение продуктивности среды при III палеопротерозойском низкоширотном оледенении (III Гуронском) 2,222 ± 0,013 млрд лет назад [181; 313; 465] подстегнуло тягу жизни к самостоятельности вообще и автотрофности в частности. Поэтому жизнь ок. 2,2 млрд лет назад овладела способностью фиксировать азот [593], что ослабило ее зависимость от стационарных природных источников этого элемента, необходимого для поддержания существования живых организмов. Здесь мы имеем дело с одним из ранних фактов повышения мобильности организмов под влиянием оледенения, точнее под воздействием вызванной им убыли продуктивности среды. Другим способом повысить мобильность организмов выступила многоклеточность. Современные бактерии способны образовывать колонии с высокоупорядоченным строением, в которых бактерии дифференцируются на клетки различных типов. Колонии некоторых миксобактерий (например Chondromyces crocatus) формируются в виде многоклеточного плодового тела со скоплениями одноклеточных спор на концах ответвлений. Это едва ли не прообраз многоклеточных растений, переживший свое время. Другие миксобактерии (например Myxococcus xanthus) образуют сферичные колонии из миллионов клеток. Такие колонии способны двигаться, захватывать и переваривать жертвы. Подобные хищные сферы в чем-то напоминают многоклеточных животных [115]. Многоклеточность последнего типа нацелена на подвижный сбор разреженной пищи, т. е. на ледниковые условия. В соответствии с такими представлениями о назначении многоклеточности первые макроскопические многоклеточные водоросли появились в палеонтологической летописи 2,110 ± 0,052 млрд лет назад ([399; 698]) в Даспортскую ледниковую эпоху (2,2–1,95 млрд лет назад), вызванную образованием следующего сверхматерика (2,1–2,0 млрд лет назад). Наступившие вслед за Даспортским оледенением условия климатического оптимума расширили экологические ниши. Это произошло потому, что мягкие условия способствовали подъему биопродуктивности среды, а ее богатство сопровождалось ростом разнообразия природных условий и, соответственно, раздвижением экониш, что в свою очередь благоприятствовало умножению и экспансии жизни. Судя по всему, простейшие организмы 1,2 млрд лет назад освоили новые для себя наземные водоемы [423]. Одновременно произошла крупная радиация (всплеск разнообразия) многоклеточных организмов, выразившаяся в разделении предков растений, грибов и животных [294; 322]; cp. [312; 780]. По молекулярно-генетическим данным, процесс продолжился отделением от животных губок, затем гребневиков, медуз и морских анемон, далее – плоских червей [312], а потом – дивергенцией (разделением) хордовых животных (к которым принадлежат и люди) и моллюсков (1,225 млрд лет назад), разделением хордовых и кольчатых червей (1,204 млрд лет назад), расхождением хордовых и членистоногих (1,173 млрд лет назад) и дивергенцией хордовых и иглокожих (1,001 млрд лет назад) [824]; cp. [294; 322; 470]. Дальнейший путь развития животных отмечен так называемой преэдиакарской биотой 980–900 млн лет назад [259, с. 220, рис. 1], которая связана с теплым эпизодом, благоприятным для умножения экониш и расцвета фауны. Вслед за распадом еще одного сверхматерика возраста 1,1–1,0 млрд лет назад наступали ледниковые периоды Гнейсе, 900–850 млн лет назад, и Стёрт, 760–740 млн лет назад, отвечавшие состоянию разреженности материков. Очередной суперконтинент сформировался 675–600 млн лет назад, что повлекло за собой низкоширотное оледенение Марино-Варангер ок. 680–606 ± 3,7/ – 2,9, 602 ± млн лет назад [313; 381; 465; 807]. Низкоширотность (тропичность) оледенений Гурон III и Варангер, определенная палеомагнитным методом (когда по намагниченности ископаемых пород рассчитывают положение магнитных полюсов в древности), объясняется дрейфом магнитного полюса Земли относительно географического полюса. В протерозое этот дрейф превосходил современный и заметно искажал регистрацию палеоширот по намагниченности ископаемых пород. Выгодность мобильности в ледниковых условиях оживила подвижных многоклеточных животных, представленных норами червей 620 млн лет назад [464; 682]. Последовавшее потепление эдиакарского периода (ок. 600–544 млн лет назад [312; 381]) создало множество экониш и вызвало дивергенцию бесчелюстных рыбообразных животных (круглоротых вроде миног и миксин) и челюстноротых (предков рыб) 599 млн лет назад [824; 479]. В результате развернулась богатая эдиакарская фауна возрастом в 565 ± 5–549/543 млн лет назад [203; 258; 381], отмеченная в финале своего расцвета молекулярно-генетической дивергенцией позвоночных животных (наших предков) 546 ± 18 млн лет назад [479; 838]. Палеозой (544-251 млн лет назад) Развившееся хищничество одело мягкотелых эдиакарцев в твердые наружные покровы [258], поэтому кембрийский период (544–510/505 млн лет назад) ознаменовался широким распространением животных с защитным наружным скелетом, а также хордовых животных, чей внутренний скелет позволял создавать в воде локомоторную волну (толкающую животное вперед [124]) и повышал спасительную подвижность. Хищники производят эффект снижения биопродуктивности среды, поскольку мешают консументам (в данном случае – нехищным потребителям) беззаботно кормиться. Поэтому усвоение последними упомянутой спасительной подвижности типологически близко реакции на падение биопродуктивности среды вследствие похолодания. Следом за жертвами в локомоции усовершенствовались и хищники, примером которых может служить подвижный нижнекембрийский Anomalocaris (45 см), напоминавший плоскую сигару с боковыми плавниками и парой хватательных придатков возле рта. Организмы, отставшие от описанной тенденции, пришли к массовому вымиранию в раннем кембрии [184; 674, с. 431–437]. Целым «пакетом» в нижнем кембрии отмечены такие животные, как членистоногие [85, с. 41; 211; 239]; ракообразные [712]; иглокожие [700]; полухордовые (век атдабан, ок. 525 млн лет назад) [237; 701; 703; 727]; оболочники [699]; хордовые (век ботом) [237]; черепноподобные (530 млн лет назад) [238; 308]; миногообразные и миксинообразные (ок. 530 млн лет назад) [702; 838]; в верхнем кембрии – конодонты (родня позвоночных; 515 млн лет назад) [669; 210; 325; 437] и другие позвоночные (514 млн лет назад) [479]. Так выглядел древнейший эволюционный взрыв в истории Земли, обусловленный теплом и высокой биопродуктивностью эдиакарского и кембрийского периодов. В следующем, ордовикском, периоде (510/505–438 млн лет назад) материки максимально отдалились друг от друга, что вызвало в позднем ордовике (458–438 млн лет назад) оледенение, условия которого обострили спрос на мобильность животных. Из числа головоногих моллюсков, появившихся в нижнем кембрии (например, наутилоидеи, потомки которых из рода Nautilus, живые ископаемые, и поныне здравствуют на юго-западе Тихого океана), особый расцвет в ордовике испытали представители подкласса эндоцератит [38, с. 356–359; 83, с. 173–204]. Их тело помещалось в длинно– и короткоконических раковинах прямых очертаний, игравших роль гидродинамических обтекателей. В раковине имелось образование – сифон, выполнявший гидростатические функции (он имелся и у других раковинных головоногих моллюсков и работал, как плавательный пузырь рыб, т. е. помогал своему обладателю менять общий удельный вес тела). Гидродинамическая раковина вместе с гидростатическим сифоном (он наполнял соответствующие камеры раковины азотом, аргоном и жидкостью, как у современного наутилуса, и тем самым регулировал удельный вес тела и его частей) делали эндоцератит идеальными пловцами, т. е. высокомобильными животными, типологически напоминающими по устройству подводные лодки на реактивной тяге. В связи с похолоданием, поощряющим мобильность, эндоцератиты расцвели, стали владыками морей и породили самых крупных среди известных раковинных беспозвоночных, до 9,5 м длиной [38, с. 356; 83, с. 191]. Казалось бы, такие успешные монстры должны были бы процветать долго, однако с концом позднеордовикского оледенения они неизбежно пришли в упадок. Представители ведущего отряда, эндоцератиды, просуществовали весь ордовик и вымерли – возможно, один род данного отряда (Humeoceras) дожил до середины силура (421 млн лет назад) [38, с. 358]. Парадокс этот вполне объясним, если учесть, что высокомобильные формы вроде эндоцератит были узко приспособлены к скромной экосреде окололедникового времени и лишь в ней были сильны, а с подъемом биопродуктивности среды в силуре перестали ей соответствовать и угасли. Прохладный ордовик был периодом радиации жизни [297], в особенности холодолюбивой – точнее, годной к низкой биопродуктивности среды. Холодолюбивые существа активно замещали ушедших в небытие теплолюбивых хозяев кембрия. Давление в пользу мобильных организмов привело на сушу наземную флору. Среди пионеров были наземные грибы (конец нижнего ордовика, ок. 460 млн лет назад), которые, по молекулярно-генетическим данным, возникли еще в конце Варангского оледенения ок. 600 млн лет назад [636], т. е. уже были испытаны суровыми условиями. Бок о бок с ними по суше распространялись наземные растения, отмеченные как в нижнем ордовике (475 млн лет назад), так и в верхнем (458–438 млн лет назад) [346, с. 481; 794]. В морях, судя по молекулярно-генетическим данным, появились подвижные лучеперые рыбы (450±35,5 млн лет назад) [86, с. 323–484; 479], а также акулы [468]. Лучеперые рыбы и акулы составляют свыше 95% современной ихтиофауны, захватившей моря благодаря кайнозойскому похолоданию. Их успех в современных условиях объясняется происхождением из позднеордовикского оледенения (см. далее). Акулы, помимо завидной подвижности, во многих случаях развили способность к яйцеживорождению и живорождению с подобием плацентарного вскармливания эмбрионов (как у холодолюбивых млекопитающих). Это является эволюционным признаком мобильности, поскольку потомство выводится не из стационарно отложенной икры, а на ходу, что повышает его шансы на распространение по экосреде. Добавим, что и яйцекладущие акулы по-своему мобильны. Они откладывают яйца (икринки) в твердой оболочке, чем уподобляются, например, рептилиям, у первых представителей которых способность откладывать яйца в твердой скорлупе послужила одной из основ сухопутной подвижности. Лучеперые рыбы и акулы не успели продвинуться в ордовике так сильно, как эндоцератиты, а потому благополучно перенесли позднеордовикское вымирание, более суровое к остальной биоте [184; 239; 836]. Объяснение парадоксального уклонения лучеперых рыб и акул от позднеордовикского вымирания благодаря отставанию от эндоцератитов мы приведем ниже (вместе с рядом аналогичных случаев). Силурийский (438–408 млн лет назад) и девонский (408–354 млн лет назад) периоды были временем сближения материков и, соответственно, эпохой прибывающего и отступающего потепления. По мере его углубления биопродуктивность среды возрастала, и суша превращалась в притягательную эконишу для консументов, потребителей органики, благо сухопутная флора к тому времени процветала уже не один миллион лет. В верхнем ордовике под влиянием отступления оледенения сушу освоили норные животные, в которых подозревают членистоногих многоножек [643]. Затем, в позднем силуре (век пржидоли, 414 млн лет назад), землю завоевали такие членистоногие, как паукообразные и непосредственно зафиксированные многоножки [440]. Позже, в нижнем девоне (век зиген, 401–394 млн лет назад), их состав пополнили насекомые [307; 483], чьи крылья размером до 2 см поначалу служили целям терморегуляции, а с повышением размера до 4 см стали применяться для полета [502]. Поскольку полет увеличивал мобильность насекомых, их крылья должны были служить органами движения в некий холодный период. Однако данных на этот счет пока нет. Обзаведясь фауной беспозвоночных, суша привлекла позвоночных хищников, которые прибыли в новую для себя экосреду в раннем верхнем девоне (век верхний фран, ок. 370 млн лет назад) [128а; 129, с. 420; 130, с. 508]; а также [246; 253; 274]. Вышедшие из пресноводных водоемов [834], первые земноводные были шестипалыми [345]. Они происходили от кистеперых рыб [334], а те – от рыб вроде Styloichthys changae (век поздний лочлов (жедин), нижний девон Китая), связывающих предков бесчелюстных рыбообразных с предками четвероногих позвоночных (земноводных, пресмыкающихся и млекопитающих) [835]. Рожденные межледниковьем, земноводные были маломобильными существами, а потому держались водоемов, привязанные к ним способом размножения (когда икра откладывается в воду, и там же развивается молодь, как у современных лягушек). Можно было бы предсказать, что, попади земноводные под давление отбора на высокомобильность, они изменили бы способ размножения на независимый от водной среды, принявшись прочесывать сушу и обзаведясь для этого непроницаемыми покровами тела взамен слизистой голой кожи, служащей для дыхания. Все это осуществили пресмыкающиеся. По мере сближения материков климат девона портился, биопродуктивность среды падала, и маломобильные организмы погружались в кризис, что выразилось в позднедевонском вымирании (рубеж веков фран/ фамен, 367 млн лет назад), которое, помимо всего прочего, вызвало гибель тропических коралловых рифов и убыль чистой биомассы [184; 239]. Это прямо отражало снижение палеотемператур (чего не переносили прикрепленные маломобильные коралловые полипы) и оскудение среды. Вслед за теплым девоном наступили каменноугольный (карбон, 354–286 млн лет назад) и пермский (286–251 млн лет назад) периоды. В это время континенты все более сближались, чтобы образовать сверхматерик Пангея (ок. 250–180 млн лет назад). Эти события вызвали пермо-карбоновое, или гондванское (по южной половине Пангеи – Гондване), оледенение (310–260 млн лет назад). Под давлением отбора на мобильность в соответствии с вышеописанным сценарием земноводные превратились в амниот (от греч. '?????? – околозародышевый пузырь), т. е. начали откладывать яйца в плотной зародышевой оболочке, позволяющей им развиваться на суше. Защищенная чешуей кожа открывала недавним амфибиям сухопутные просторы. Появились пресмыкающиеся (век бригант (визей), нижний карбон, ок. 338 млн лет назад) [716]. На протяжении пермо-карбонового оледенения пресмыкающиеся сохраняли четвероногость, которая обеспечивала им максимальную мобильность. Ошибочно думать, будто ранние пресмыкающиеся не были в состоянии усвоить какую-либо другую локомоцию. Напротив, в их среде появилась интересная ящерицеобразная рептилия Eudibamus cursoris (из числа Bolosauridae, ранняя нижняя пермь, век ассель, 290 млн лет назад) [187]. Она напоминала ряд современных ящериц малопродуктивных, открытых пространств, бегающих рысью, переходящей в бег на двух задних ногах (такова, например, австралийская плащеносная ящерица), что отличается от энергосберегающей шагающей двуногой локомоции динозавро-человеческого типа. Отсюда следует, что E. сursoris был порожден оледенением и не имел ничего общего с двуногими динозаврами и людьми (в смыле конвергентной, внешне похожей эволюции), но подтверждал способность ранних рептилий разнообразить свою походку. Здесь необходимо остановиться на способах локомоции (передвижения), присущих высшим наземным позвоночным. За вычетом промежуточных вариантов вроде бега E. cursoris или продвинутых кроличье-лягушачьих прыжков кенгуру, тушканчиков и им подобных животных существуют два основных типа локомоции: четвероногая, как у подавляющего большинства современных млекопитающих и «холодолюбивых» динозавров, и шагающая двуногая, как у «теплолюбивых» динозавров, людей и некоторых других существ. Четвероногость весьма расточительна энергетически, сопряжена со значительным сцеплением животного с субстратом (из-за четырех конечностей), однако обеспечивает ему исключительную выносливость, проходимость, значительные скорости и дистанции передвижения. Подобная адаптация крайне полезна в прохладных условиях низкой биопродуктивности среды, когда скудную пищу приходится собирать с больших пространств. В противоположность этому шагающая двуногость весьма выгодна, экономна энергетически, не сопряжена с большим сцеплением тела с субстратом (благодаря опоре на две конечности), однако не дает своему обладателю выносливости и не позволяет осваивать значительные территории. Такой способ передвижения выгоден в теплые эпохи высокой биопродуктивности среды, когда пища богата, скучена в пространстве и не требует больших путешествий для своей добычи. При этом двуногое существо очень выигрывает, экономя энергию на передвижении. С конкретными примерами такого рода мы познакомимся ниже. Пермокарбоновое оледенение отмечено еще двумя перспективными достижениями биосферы. По молекулярно-генетическим данным, в это время возникли цветковые (покрытосеменные) растения. В конце нижнего карбона (век намюр, 319 ± 35 млн лет назад) они разделились на однодольные и двудольные, а в перми (276 ± 33 млн лет назад) двудольные растения претерпели радиацию [545]. Покрытосеменные растения были родственны растениям из рода Glossopteris, преобладавшему в перми и отличавшемуся простым способом размножения, как у цикадовых и гинкго [595]. Происходя из холодного ордовика, все наземные растения имели мобильные средства размножения (споры, семена) и совершенствовались в этой части. Возникнув в холодном карбоне, цветковые растения сделали здесь очередной шаг вперед: благодаря различным приспособлениям их семена распространялись на большие расстояния лучше, чем у голосеменных (например хвойных), хотя последние уступили преобладающее положение не сразу [88, с. 21–382]. Другое достижение касается холодолюбивой фауны, в рамках которой пресмыкающиеся котилозавры породили в верхнем карбоне звероподобных рептилий, господствовавших в перми, постепенно убывающих в триасе, вымирающих в средней юре, однако отмеченных в поздней юре и даже в позднем палеоцене (век ранний тиффан, ок. 60–57,3 млн лет назад) [25, с. 110–117; 87, с. 230–298; 250; 328; 655, с. 173–186]. От этих четвероногих рептилий в перми произошли зверозубые пресмыкающиеся – чрезвычайно прогрессивные существа: некоторые, по-видимому, стали теплокровными, покрылись шерстью и усвоили разнозубость. Зубы прочих пресмыкающихся обычно однородны (за вычетом динозавра гетеродонтозавра из позднего триаса или ранней юры (ок. 201 млн лет назад), у которого самцы имели клыки [37, с. 46–47]; гетеродонтность динозавра Eoraptor lunensis из верхнего триаса (ок. 230 млн лет назад) далека от разнообразия зверозубых рептилий и млекопитающих, cp. [689, с. 64, рис. 1]). У продвинутых же зверозубых пресмыкающихся зубы дифференцированы на резцы, клыки и коренные (как у их потомков – млекопитающих). Это требовалось для эффективного использования небогатой и разреженной пищи ледниковья. Казалось бы, перед прогрессивными зверозубыми рептилиями открыт весь мир. Между тем, их постепенно смело с эволюционной сцены. С концом пермского похолодания грянуло позднепермское вымирание, неблагосклонное к мобильным организмам. Однако оно лишь потеснило цветковые растения и зверозубых рептилий. Это вымирание осуществлялось в две стадии: гваделупа, ок. 256 млн лет назад, истребившая 58% родов морских организмов, и татарий, ок. 251 млн лет назад, уничтоживший 67% родов морских организмов [723; 825 a]; cp. [35, с. 87, рис. 2; 184; 239; 311; 471; 639; 674, с. 437–439; 802]. Сухопутная биота понесла сходные потери, отвечающие эволюционно-экологической направленности естественного отбора, связанного с потеплением. Закономерно были утрачены небольшие подвижные рыбоеды и насекомоядные существа. Их участь разделили средние и крупные травоядные, привыкшие собирать скудные корма с обширных пространств. Высшие хищники потянулись следом. Даже спустя 15 млн лет этот урон среди пресмыкающихся еще не восполнился [185]. Легко понять, что избирательное двухступенчатое пермо-триасовое вымирание подчинялось определенной эволюционной логике, а не уничтожало живых существ без разбора, как свойственно стихийным бедствиям вроде падений астероидов или извержений вулканов. Как и в случае с мелтретичным вымиранием (см. далее), углубление исследований рисует эти биокатастрофы все менее окказионально (т. е. все менее случайно). Мезозой (251-65 млн лет назад) Наступил триасовый период (251–201 млн лет назад), начало которого ознаменовалось «метановым потеплением» (вследствие парникового эффекта от вулканических газов, включая метан) [742] и морской трансгрессией (подъемом уровня Мирового океана) [74, с. 219, рис. 7.1; 401]. Естественный отбор принялся поощрять маломобильных животных. Еще в верхней перми от рептилий-эозухий, близких прогрессивным котилозаврам-капторинам и низшим зверообразным пеликозаврам (заложившим черты молекулярно-генетического родства своих потомков-млекопитающих и людей с потомками текодонтов, т. е. птицами), произошли пресмыкающиеся текодонты (псевдозухии). Под влиянием потепления и связанного с ним роста биопродуктивности среды, они усвоили экономичную шагающую двуногость человеческого типа, длительное время процветали, однако не перенесли похолодания (падения биопродуктивности среды) в конце триасового периода и вымерли. Однако до того, в середине триаса, морская трансгрессия и потепление обострили давление отбора в пользу маломобильных животных. В результате от текодонтов монофилетично (из одного корня) произошли двуногие динозавры двух отрядов (ящеротазовые и птицетазовые), 227,8 ± 0,3 млн лет назад (см. рис. 3) [87, с. 494, 525; 271; 324; 329; 653; 655, с. 138–141; 688–690]. Судя по своим поздним потомкам мелового периода и подобно ближайшим родичам, птицам и крокодилам, динозавры имели четырехкамерное сердце. Аналогично птицам они были покрыты перьями [148; 441; 747; 827–829], возможно, отличались постоянной температурой тела (гомеотермией) [173]; cp. [660] и даже спали в птичьей позе [826]. Они были сходны с птицами в особенностях роста челюсти [172], но расходились с ними в скоростях общего роста [310], что можно объяснить повышенным метаболизмом (обменом веществ) у последних вследствие летающего образа жизни. Кроме того, двуногим динозаврам, даже их верхнемеловому венцу, хищному колоссу тираннозавру, были присущи маломобильность [430] и равнозубость, ориентированная на расточительное потребление богатой и скученной пищи, – то и другое гармонировало с высокой биопродуктивностью среды. Длительное триасовое потепление раздвинуло существующие экониши и вовлекло в них маломобильных существ, что привело пресмыкающихся в водную среду и дало известных ихтиозавров (средний триас – поздний мел), плезиозавров (тогда же) и других водных ящеров [25, с. 18, 22, 33]. Похолодание позднего триаса (норий-рэтийская морская регрессия, падение уровня Мирового океана) привело к упадку и частичному вымиранию маломобильных ихтиозавров. Выжили те их формы, которые лучше освоили живорождение, являющееся вариантом неявной мобильности. То же похолодание отпочковало от динозавров подвижных летающих птиц, засвидетельствованных спорными остатками Protoavis texensis («первоптицы из Техаса», поздний триас, ок. 225 млн лет назад [141]; cp. [240; 326]), ископаемыми птичьими следами (поздний триас, века норий-рэт, св. 212,5 ± 7 млн лет назад [565]) и синхронным молекулярно-генетическим расхождением птиц с крокодилами (222 ± 52 млн лет назад [479]), что, впрочем, не исключает обособления предков птиц от крокодилов еще в составе динозавров (т. е. сначала от крокодилов отделились динозавры, а впоследствии, много позже, от динозавров обособились птицы вроде археоптерикса, которые положили начало современной авиафауне, что представляется нам сомнительным, см. ниже).  Рис. 3. Ставрикозавр (Staurikosaurus pricei, «ящер созвездия Южного Креста») из семейства ставрикозаврид подотряда теропод (звероногов) отряда заурисхий (ящеротазовых динозавров) был ловким двуногим хищником и охотился в среднем триасе, не подозревая, что в конце этого периода его отсталые родичи прозауроподы встанут на четвереньки и дадут начало крупнейшим наземным животным планеты, зауроподам. [37, с. 30–31] Позднетриасовое вымирание (210 млн лет назад [184; 239; 518]) затронуло малоподвижных животных, но оживило четвероногих, что выразилось в оформлении млекопитающих как класса позвоночных животных (век норий, св. 200 млн лет назад [330]). Однако затем рэтийская регрессия ненадолго сменилась парниковым потеплением на 3–4° вследствие роста вулканического диоксида углерода 205,7 ± 4 млн лет назад [560], что спровоцировало вымирание мегафлоры, ориентированной на падение биопродуктивности среды. Наступивший юрский период (201–144,2 млн лет назад) открылся очередной регрессией (похолоданием) и потряс двуногих динозавров – произошло их ограниченное нижнеюрское вымирание [239, с. 49]. Если целурозавры и карнозавры всегда оставались удачливыми, а потому эволюционно негибкими двуногами, то неуклюжие двуногие прозауроподы были готовы ко всему новому (о причинах чего см. далее) и охотно перешли к четвероногой локомоции, превратившись в гигантских зауропод (нижняя юра, век поздний лейас – верхний мел) (см. рис. 4, 5). Отметим впечатляющую парадоксальность эволюционного процесса: предки динозавров (текодонты) сначала «встают» с четырех ног на две, порождают успешных двуногих динозавров, а затем вновь «опускаются» на четыре конечности (как произошло в случае с зауроподами и некоторыми другими динозаврами, см. далее). В рамках нашей версии эволюционной направленности этот нонсенс находит естественное объяснение – климато-экологическое. Возможно, обсуждаемое нижнеюрское похолодание развило у гетеродонтозавров разнозубость, являющуюся адаптацией к низкой биопродуктивности среды, свойственной похолоданиям. После метанового потепления (ок.183 млн лет назад [742]) в середине средней юры (век поздний байос) палеотемпературы упали с наступлением антарктического микрооледенения, о чем свидетельствуют тиллиты (ископаемые морены, ледниковые отложения) юрского времени в Антарктиде [42, с. 33; 74, с. 220, 222], однако затем тепло стало нарастать к максимуму трансгрессии в середине верхней юры (конец века кимериджа). Средне-верхнеюрское потепление пагубно сказалось на четвероногих звероподобных рептилиях, которые почти полностью вымерли, оставив по себе молодых отпрысков, млекопитающих. То же самое произошло с четвероногими зауроподами, испытавшими с потеплением первый эпизод вымирания [25, с. 56]. Их судьбу повторили четвероногие птицетазовые динозавры сцелидозавры из ранней юры, когда их четвероногость была закономерна. Во второй половине юрского периода они дали четвероногих же стегозавров (середина юры – ранний мел), а через сирмозавров (конец раннего мела) – панцирных анкилозавров (ранний – поздний мел) [25, с. 79]. Иными словами, эволюция этих существ шла как бы «ракетным способом», когда в моменты потеплений отбрасываются «отработанные» (устаревшие и вымирающие) ступени.  Рис. 4. Платеозавр (Plateosaurus engelhardti, плоский ящер Энгельгардта) из семейства платеозаврид инфраотряда прозауропод (проящероногов) из подотряда теропод группы зауроподоморфов (ящероногоподобных) отряда заурисхий. Это неуклюжее двуногое животное питалось растительностью, но происходило от совершенных двуногих динозавров среднего триасового периода, хищных теропод. В позднем триасе под влиянием похолодания и падения биопродуктивности среды прозауроподы стали возвращаться к четвероногому способу передвижения, свойственному далеким «холодолюбивым» предкам двуногих текодонтов и динозавров [37, с. 34–35] Нижнеюрское похолодание с его давлением в пользу мобильности выщепило из среды архозавров (родственников динозавров) летающих ящеров птерозавров, покрытых шерстью. Они просуществовали до верхнего мела и разделили судьбу динозавровой биоты [25, с. 106–109]. Их вымирание объясняется тем, что, в отличие от сноровистых, высокоподвижных птиц современного типа, они были планирующими, энергосберегающими созданиями, а не активными летунами, т. е. характеризовались ограниченной мобильностью [25, с. 106; 87, с. 593], в то время как маастрихтское (конца мела) и палеоценовое (после мела) похолодания требовали от животных более сильно выраженной подвижности.  Рис. 5. Барапазавр (Barapasaurus tagorsi, большеногий ящер) из подотряда зауропод (ящероногов) группы зауроподоморфов отряда заурисхий. Под влиянием ухудшения климата и падения биопродуктивности среды неуклюжие двуногие прозауроподы породили в раннем юрском периоде совершенных четвероногих зауропод. Они процветали и достигли размеров, рекордных для наземных животных [37, с. 42–43] В поздней юре (150,7–144,2 млн лет назад) максимум трансгрессии сменился регрессией и похолоданием, что возобновило селекцию на подвижность и побудило динозавров вторично породить птицеподобных существ – в частности, известного археоптерикса [240, с. 349], который строением зрительной и слуховой коры головного мозга действительно напоминал современных птиц [136], что явилось результатом конвергенции, параллельной эволюции. Будучи предком энантиорнитинов, или «противоположных птиц» (от последовательности совмещения костей стопы в ходе развития, противоположной современным птицам), археоптерикс напоминал миниатюрного велоцираптора или дромеозавра [697], тоже пернатых динозавров, и отличался от протоависа, предка орнитуринов, давших современных птиц. Динозавров, археоптерикса и энантиорнитинов объединяли примитивные признаки эктотермности, т. е. непостоянной температуры тела [660]. Меловой период (144,2–65 млн лет назад) ознаменовался нижнемеловой регрессией (век валанжин, 130 млн лет назад) и продолжением отбора на мобильность. В это время развивались упомянутые тупиковые энантиорнитины и перспективные орнитурины, в среде которых началась дивергенция современных птиц (средний нижний мел, век апт, 122,9 ± 0,3, 121,2 ± 0,3–121,1 ± 0,2 млн лет назад) [260; 747]. За тот же временной интервал пернатые динозавры породили еще одно птицеобразное создание – четырехкрылого Microraptor gui из Китая [829], все четыре конечности которого годились для планирования, повышая подвижность. Среди современников этого маленького дромеозавра – китайских птиц-энантиорнитинов – также встречался четырехкрылый вид [833], так что данная эволюционная тенденция проявлялась неоднократно. Если зауроподы откликнулись на падение биопродуктивности среды мобильной четвероногостью, то орнитоподы, ящеротазовые четвероноги (игуанодоны) и двуноги (утконосые динозавры-гадрозавры, верхняя юра – верхний мел) отреагировали иначе: они освоили умение жевать, как млекопитающие [596], что повышало эффективность потребления редкой пищи соответственно похолоданию. В этом отношении названные представители динозаврового племени принципиально отличались от прочих архозавров, т. е. других динозавров, а также крокодилов. Последние, кстати, пережили роковой меловой период в силу того обстоятельства, что являлись ловкими наземными бегунами-четвероногами, т. е. мобильными животными, что открывало им путь в холодный кайнозой, где они, впрочем, вскоре «замерзли» и перешли к водному образу жизни в тропических и субтропических реках, не выдержав конкуренции с крепнущими наземными четвероногими млекопитающими. Рубеж нижнего-верхнего мела (105 млн лет назад) прошел с трансгрессией, а века турон – коньяк (92–86 млн лет назад) подверглись вулканическому диоксид-углеродному потеплению [750]. В конце мела, в маастрихте (71–66 млн лет назад) регрессия сопровождалась заметным похолоданием [526], когда тропические температуры в океанах упали ниже современных [288]. К этому времени изнеженные теплым и обильным мезозоем (251–65 млн лет назад) динозавры находились на вершине своего расцвета, о чем свидетельствует сам факт существования крупнейшего наземного хищника – королевского тираннозавра (76–65 млн лет назад) (см. рис. 6). Сопряженная с ним мегафауна стала совершенной и высокоспециализированной, а потому могла лишь углубляться в аппетитах, но никак не ограничивать себя, чего требовала оскудевающая обстановка. Вымирание динозавров происходило постепенно и пропорционально прогрессу млекопитающих (по модели цветковых растений и костистых рыб, см. далее). В веке кампан (76–73 млн лет назад) на пик расцвета динозавров приходилось 30 их родов. С приближением маастрихтского похолодания (71–66 млн лет назад) гордые животные ощутили эволюционно-экологическое затруднение: они любили много потреблять и мало двигаться, а теперь среда благоволила к сноровистым аскетам типа четвероногих млекопитающих.  Рис. 6. Тираннозавр (Tyrannosaurus rex, королевский ящер-тиран) из семейства тираннозаврид инфраотряда карнозавров (хищных ящеров) подотряда теропод отряда заурисхий. Это был крупнейший наземный хищник всех времен, чуть уступавший лишь хищному гигантозавру начала верхнего мела (ок. 100 млн лет назад), который, будучи южноамериканцем, закономерно превосходил массой североамериканского собрата, как свойственно рептилиям с приближением к экватору (у млекопитающих – наоборот). Тираннозавр был совершенным хищником (и падальщиком, как современные львы), узко приспособленным к среде, богатой пищей. С ухудшением климата и падением биопродуктивности среды королевский ящер угас и вымер вместе с подавляющим большинством остальных динозавров. По современным данным, он был оперенным существом [37, с. 98–99] За весь маастрихт и две трети дания (72–65,3 млн лет назад) число родов динозавров упало до 12. Затем вымирание ускорилось, и за период 65,3–64, 96 млн лет назад прошло пять убывающих стадий: 12–12–11–10–7 родов уходящих динозавров. О том, что биосфера отреагировала на коварный удар иридиевого астероида 65 млн лет назад, не может быть и речи. Параллельно со ступенчатым падением динозавров шел уверенный прогресс копытных млекопитающих, представленных в тех же временных точках 0–1 – 3–5 – 8 родами. Так что за 40 тыс. лет, истекших с момента падения астероида, млекопитающие зримо окрепли, а динозавры еще не вымерли окончательно, являя редкий случай послемеловых, палеоценовых динозавров, останки которых обнаружены выше мелтретичной иридиевой аномалии, оставленной упавшим болидом [714; 713]; cp. [222; 642; 696]. Приведенные количественные данные по динозаврам и копытным млекопитающим за период 65,3–64,96 млн лет назад происходят из формации Хелл Крик в Монтане, США (северо-запад страны). Монтана лежит на том же континенте, что и мексиканский кратер Чикхулуб, оставленный ударным небесным телом 65 млн лет назад. Монтанская биота совершенно не отреагировала на это событие: млекопитающие набирали эволюционные обороты, а динозавры сокращались в разнообразии в соответствии с тенденцией, наметившейся за 7 млн лет до вторжения астероида. Не избежали их судьбы и обитатели других сред. Морские пресмыкающиеся (ихтиозавры, плезиозавры, мозозавры и пр.), страдавшие низкой мобильностью [35, с. 205–206], вымерли в верхнем меле: более подвижные ихтиозавры – в его теплом начале, остальные – ближе к холодному концу [25, с. 18, 33]. Господствовавших на море раковинных головоногих моллюсков аммонитов и белемнитов постигла та же участь. Показательна в этой связи эволюционная история теплолюбивых аммоноидей (нижний девон – верхний мел) [83, с. 243–425; 84, 15–144]. Появившись в начале раннего девона (век жедин, 408–401 млн лет назад), они продолжили линию других головоногих моллюсков – бактритов, имевших прямую или слабосогнутую раковину, т. е. являвшихся более-менее хорошими пловцами. Однако «возниковение аммоноидей было связано с образованием вначале согнутой, а затем криоконовой раковины, состоявшей из несоприкасающихся оборотов, свернутых в одной плоскости» [38, с. 376]. Это делало аммоноидей слабыми пловцами и приводило в соответствие с теплым, высокобиопродуктивным девоном, когда была выгодна умеренная мобильность. В эпоху пермо-карбонового оледенения ситуация изменилась, и малоподвижные аммоноидеи закономерно отступили в эволюционную тень, поскольку оледенение благоприятствовало более энергичным созданиям. Зато теплый мезозой востребовал их качества, и в кампане – маастрихте (76–66 млн лет назад) они выдвинули род Pachydiscus, у представителей которого свернутая раковина достигала рекордных 2 м в диаметре [84, с. 111]. Напомним, что скоростное плавание обеспечивала прямая раковина (см. выше об эндоцератитах). Свернутая же в диск раковина, наоборот, скоростей не давала. При этом под влиянием климатических колебаний в меле аммониты то пытались слепо распрямить раковину, чтобы повысить свою мобильность и попасть в унисон с похолоданиями, то сворачивали ее обратно, чтобы убавить подвижность и совпасть с потеплениями. В силу закона Долло о необратимости эволюции эти попытки были обречены на провал и лишь наводняли меловые моря уродливыми созданиями. Избирательность верхнемелового вымирания прослеживается в судьбе двустворчатых моллюсков. Их прикрепленные формы вымирали заметно интенсивнее, нежели свободноживущие [434]. Это объясняется тем, что прохладное дыхание наступающего кайнозоя готовило перспективу лишь подвижным существам – в данном случае свободноживущим двустворчатым моллюскам. С мезозойскими воздухоплавателями происходило то же самое. Отряды подвижных современных птиц дивергировали еще при похолодании в раннем меле, и 22 их филетические линии успешно пересекли мелтретичную границу, не побоявшись зловещего астероида [260]; ср. [252]. Однако сам факт биологической принадлежности к современным птицам отнюдь ничего не значил. В частности, могучие (более метра в высоту) нелетающие гесперорнисы были водоплавающими птицами и в силу своей маломобильности не перенесли верхнемелового отбора. Маломобильным планирующим птерозаврам тоже не повезло, равно как и «противоположным птицам» энантиорнитинам, которые летали хуже тогдашних орнитуринов, предков неорнитов, нынешних птиц [697]. Между тем, финал мелового периода не был эпохой тотального вымирания [35, с. 89, рис. 3; 184; 239; 394; 407; 674, с. 439–441]; cp. [444]. Эволюционная история меловой биоты выглядела как пара сообщающихся сосудов, в одной из емкостей которых убывали многочисленные маломобильные господа мезозоя (динозавры, птерозавры, ихтиозавры, плезиозавры, аммониты, белемниты и др.), а в другой прибывали скромные высокомобильные господа будущего кайнозоя (покрытосеменные растения, млекопитающие, современные птицы, костистые рыбы, акулы, внутреннераковинные головоногие моллюски и др.). Будучи родом из пермо-карбонового оледенения (см. выше, а также [284; 407; 638]), цветковые растения положительно отреагировали на ухудшающуюся климатическую обстановку нижнего мела и в середине мела (кон. века альба, 105 млн лет назад) произвели революцию флоры. С верхнего мела к ним перешло флористическое господство, которое укрепилось в кайнозое и наиболее ярко представлено сейчас [88, с. 423]. Через своих рептильных звероподобных предков млекопитающие тоже ведут начало из пермо-карбонового оледенения. Оттуда происходит их четвероногая мобильность и эффективная разнозубость, выгодные в условиях скудной биопродуктивности среды. Как класс, они выщепились в холодное время норий-рэтийской регрессии. В поздней юре или раннем меле из их пула выделились однопроходные звери (ок. 144,2 млн лет назад) [149; 466]. По молекулярно-генетическим данным, то же самое произошло с сумчатыми млекопитающими еще раньше (в средней юре, 173 ± 12,3 млн лет назад) [479], хотя палеонтологически они известны лишь с позднего мела (84–65 млн лет назад) [183]. По молекулярно-генетическим данным, пять линий высших плацентарных млекопитающих обособились св. 100 млн лет назад, т. е. в начале верхнего мела (в веке сеномане, 105–92 млн лет назад), и большинство их современных отрядов появилось до мелтретичной границы; палеонтологически плацентарные млекопитающие отмечены 85 млн лет назад [479]; а также [651; 655, с. 187–310]. Радиация сумчатых и плацентарных млекопитающих связана с теплыми периодами юры и мела, когда подавляющие их четвероногие динозавры испытывали эволюционную депрессию, и вакансии в соответствующих эконишах доставались млекопитающим. Последние были преимущественно мелкими формами с черно-белым зрением, поскольку вели скрытный, ночной образ жизни. Однако среди них встречались и более выразительные представители. В раннем меле Китая, 139–128 млн лет назад, существовал хищник из числа млекопитающих Repenomomus giganticus, достигавший свыше метра в длину и 12–14 кг массы, что позволяло ему успешно охотиться на детенышей динозавров (в частности, на пситтакозавров, один из которых сохранился у него в брюхе [425]). Млекопитающие, подвижные зверьки, настойчиво прогрессировали и, подобно птицам, уверенно пересекли мел-палеоценовую границу. Параллельно цветковым растениям, млекопитающим и птицам меловую экосреду завоевывали костистые рыбы. Являясь отпрысками лучеперых рыб, предки которых зародились в период ордовикского оледенения, костистые пловцы были сильны в прохладной, малопродуктивной среде, поскольку лучше других рыбообразных создавали осевым скелетом двигательную локомоторную волну [124]. Недаром их современный рекордсмен, меч-рыба, развивает огромную скорость до 130 км/ч, превосходя по этой части наземного гепарда. Непреодолимо прогрессируя с раннемелового похолодания, они возобладали в морях с верхнего мела, легко пересекли мелтретичную границу и ныне составляют 95% ихтиофауны [86, с. 334; 655, с. 61–71]. Эта победа облегчалась малой мобильностью прочих мезозойских рыб [35, с. 205]. Эволюционная история акул, также имеющих ордовикские корни и отличающихся высокими ходовыми качествами, развивалась по образцу костистых рыб [86, с. 196–237] и тоже отмечена рекордами в кайнозое (см. ниже). Наконец, безуспешная борьба за жизнь медлительных аммонитов и белемнитов закончилась вытеснением их подвижными внутреннераковинными головоногими моллюсками: десятиногими кальмарами и каракатицами, а также и восьминогими осьминогами [35, с. 187; 84, с. 145–178]. С падением общеглобальной биопродуктивности среды разнообразие жизни на планете убывает, и проявления эволюционной направленности теряют разнообразие. Масштабы организмов тоже мельчают. Поэтому господствующие наземные животные мезозоя были крупнее кайнозойских, а раннекайнозойские – внушительнее позднекайнозойских современников нарастающего похолодания. Соответственно, примеры нашей версии эволюции для кайнозоя относительно скромны. Кайнозой (65-0 млн лет назад) В палеоцене (65–55 млн лет назад) высокомобильные организмы мелового периода продолжают разнообразиться, не меняя своей эволюционной направленности, поскольку вся эпоха отмечена регрессией и похолоданием. В начале эоцена (55–36 млн лет назад) началось метановое потепление [742], и регрессия сменилась трансгрессией. Следовало бы ожидать, что в установившихся теплых условиях появятся маломобильные формы мезозойского типа. В соответствии с этим ожиданием в среднем эоцене распространяются уцелевшие потомки вымерших меловых белемнитов – байанотеутиды [35, с. 189–190; 38, с. 382, 388, рис. 217; 84, с. 161–162]. Тогда же возникают не связанные общностью происхождения нелетающие, бегающие птицы Ratidae, своего рода конвергенты (т. е. внешние подобия) двуногих динозавров: страусоподобные элеутерорнисы (средний эоцен, век лютет) и эпиорнисовые псамморнисы (эоцен) [35, с. 245–246]. Наконец, в раннем среднем эоцене Германии (50–48 млн лет назад) обнаруживается двуногое бегающее млекопитающее Leptictidium nasutum, подлинный конвергент двуногих динозавров со свойственным им и людям прямохождением (см. рис. 7) [118, с. 34]. Эоценовое потепление повысило биопродуктивность среды и раздвинуло рамки экониш, привлекая в них новых обитателей. В соответствии с этой новацией сухопутные парнокопытные произвели водных китообразных в нижне-среднем эоцене (ок. 52 млн лет назад [752]) или в конце среднего эоцена (в лютете, 47 млн лет назад [354]). В гармонии со своим происхождением в теплую эпоху китообразные (зубатые киты) развили чуждую млекопитающим равнозубость динозаврового типа. В олигоцене (36–24 млн лет назад) трансгрессия прекращается и сменяется регрессией во второй половине эпохи. В связи с похолоданием высокомобильные четвероногие млекопитающие испытывают эволюционный подъем и порождают мегафауну, украшением которой является гигантский безрогий носорог Indricotherium transouralicum – самое крупное и высокое наземное млекопитающее, до 5 м высотой (средний олигоцен). 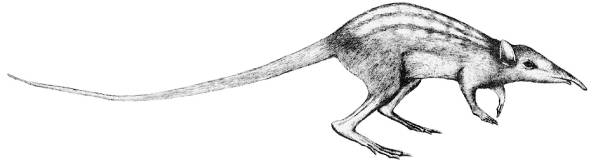 Рис. 7. Легкобег носатый (Leptictidium nasutum) возник в эпоху раннего среднего эоцена на островах нынешней Германии 50–48 млн лет назад, когда из-за потепления поднялась биопродуктивность среды, и естественный отбор дал шанс появлению маломобильных двуногих животных. «Носач» бегал на двух ногах, как двуногие динозавры и люди, а потому являлся их эволюционным конвергентом (подобием), таким же «теплолюбивым», как и они [118, с. 34] В миоцене (24–5,3 млн лет назад) ранне-среднемиоценовое потепление (24–13 млн лет назад) положило конец олигоценовой мегафауне. Зато пробившийся отбор на маломобильность вызвал к жизни двуногую обезьяну «перелоапитека каталонского» (Испания, 13 млн лет назад), положившего начало целой плеяде двуногих гоминин (см. разд. 2. 4). Происходя от аборигенов высокобиопродуктивных тропических лесов, они органично несли в себе мезозойскую ориентацию на маломобильность и прожорливость, что в сочетании с эволюционной отсталостью создало гоминин, перспективных господ будущего, ставших лидерами именно в силу своей отсталости. В плиоцене (5,3–1,6 млн лет назад) начало эпохи ознаменовалось трансгрессией. Затем последовал ряд гляциалов (оледенений), интергляциалов (межледниковий), стадиалов (пиков оледенений) и интерстадиалов (смягчений оледенений) выраженной ледниковой эпохи. Ее «лицом» можно назвать тридцатиметровую белую акулу Carcharodon megalodon, вымерщую в среднем плиоцене (3,3 млн лет назад), когда интергляциальная трансгрессия Астий вызвала ограниченное вымирание в среде мобильной биоты; cp. [680]. Наконец, в плейстоцене (1,6–0,0117 млн лет назад) развилась богатая приледниковая мамонтовая фауна, вымершая с началом голоценового потепления 11700 календарных лет назад, или 10200 лет назад по радиоуглероду 14С (поясним, что вследствие колебаний содержания тяжелого изотопа углерода 14С в древней атмосфере радиоуглеродные даты исследуемых образцов оказываются моложе своего реального, календарного возраста, который можно установить, например, методом дендрохронологии, когда высчитываются годичные кольца очень старых деревьев, до 8000 лет возраста, после чего итог сравнивается с радиоуглеродной оценкой; получается, например, что календарной дате 4650 лет назад отвечает радиоуглеродная дата 4050 лет назад). Подобно динозаврам, мамонты и их спутники вымерли не сразу. Некоторые виды жили дольше: островные карликовые мамонты острова Сен-Пол в Беринговом море, 7908 ± 100 лет назад [391], островов Санта-Барбара в Калифорнии [480, с. 213], острова Врангеля на границе Восточно-Сибирского и Чукотского морей, 7620–3730 лет назад [515; 768]; большерогий олень в Западной Сибири, 6900 14C, или 7700 календарных лет назад [736]; возможно, шерстистый носорог в Чувашии, 921/922 гг. н. э. [54, с. 61–66, 139–140; 91, с. 72–74]. «Лицом» нынешней голоценовой эпохи – межледниковой, однако все еще не свободной ото льдов – можно считать гигантского кальмара Architeuthis longimana 19 м длиной (возможны тридцатиметровые особи), продолжающего линию внутреннераковинных головоногих моллюсков, которые сменили аммонитов в конце мелового периода. Относительно упомянутых вымираний существуют неокатастрофические взгляды, касающиеся в первую очередь пермо-триасовых и мел-третичных событий. Согласно одному из них, массовые вымирания вызваны вулканической деятельностью планеты, которая обескислороживает океаны, отравляя их. Эти несчастья происходят с периодичностью в ~ 32 млн лет [632]. Они ответственны за массовую гибель биосферы на границе перми и триаса [227; 637; 639], а также мела и палеоцена [227; 598]; cp. [361, с. 368]. Показано, однако, что обескислороживание океанов господствовало на Земле на протяжении 250 млн лет в позднем неопротерозое ок. 800–543 млн лет назад [471; 802], однако на тогдашней жизни (например на морской эдиакарской фауне) это заметно не сказалось. Показано также, что обескислороживание океанов в самом начале триаса явилось следствием предшествующего позднепермского вымирания (кризиса радиолярий, морских микроорганизмов) [633], а никак не его причиной. Наконец, показано, что обескислороживание океанов имело место и в конце нижнего мела (в альбе, ок. 112 млн лет назад) [481], что не помешало костистым рыбам, акулам и прочим наследникам мезозоя уверенно набирать эволюционные обороты. В свете указанных расхождений с реальностью вулканическая гипотеза не может быть принята. Согласно другим представлениям, Землю бомбардируют космические тела (астероиды и кометы), удары которых поднимают в атмосферу взрывной материал, затеняют им планету и создают эффект «ядерной зимы», которая умерщвляет биосферу. О падениях небесных объектов говорят геологические прослойки редкого металла иридия, встречающегося в метеоритном веществе, как, впрочем, и в земных породах, изверженных из мантии в результате вулканической деятельности. Последнее обстоятельство заставляет исключить ископаемую иридиевую аномалию, например ордовик-силурийской границы, из числа космогенных [804]. В разведанной истории Земли насчитывают до 140 болидовых ударов [347]; cp. [361, с. 368]. Среди них наиболее известны следующие: • поздний плиоцен, ок. 2,15 млн лет назад [347] или ок. 2,3 млн лет назад [482] (астероид св. 1 км в поперечнике); • средний плиоцен, 3,3 млн лет назад [680]; • поздний эоцен, 35,7 ± 0,2 млн лет назад [199]; • ранний эоцен, ок. 50 млн лет назад [347]; • мелтретичный, 65 млн лет назад [137; 138; 192; 195; 378; 415; 433; 461–463; 477; 554; 558; 561; 563; 649; 743; 745]; болид вызвал глобальные лесные пожары [414; 771]; • поздняя юра, 140 ± 20 млн лет назад [378]; • триас-юрский [603]; • поздний триас, 211 (220–200) млн лет назад [518, с. 46]; 5 кратеров, век норий, 214 млн лет назад [721; 786]. • пермо-триасовый [175]; (251,4 млн лет назад) [180]; cp. [311]. • верхний девон, рубеж веков фран-фамен, 367 млн лет назад [247]. • средний девон, ок. 380 млн лет назад [305]. • поздний докембрий, ок. 600 млн лет назад [365; 366; 806]. • ранний протерозой, 1,85 млрд лет назад [179; 588]. • верхний ранний архей, 3,470 ± 0,002 млрд лет назад [226]. В настоящее время доказано, что после триас-юрской иридиевой аномалии, свидетельствующей о падении болида, тогдашние динозавры отнюдь не вымерли, а, наоборот, расцвели менее чем через 10 тыс. лет после грозного события [603]. Выше мы привели факты, указывающие на то, что позднепермское вымирание состояло из двух крупных стадий, разделенных 5 млн лет, так что его космический источник нереален. Приведены также факты, рисующие позднемеловое вымирание как избирательное событие в духе дарвиновской теории естественного отбора – его космический источник излишен. Кроме того, хорошо известно, что с ранней юры до позднего мела холодные высокие широты Земли населялись полярными динозаврами. Они известны из ранней юры (век плинсбах) Антарктиды [397], из раннего мела (века валанжин – альб, 130–105 млн лет назад) тогдашней заполярной Австралии [644; 142], из позднего мела (века поздний кампан – ранний маастрихт, 76–66 млн лет назад) тогдашней заполярной Аляски [215]. Хорошо приспособленные к продолжительному холоду и тьме полярной ночи, они бы встретили «ядерную зиму» с энтузиазмом. Приведенные соображения делают биологическую сторону космической гипотезы неприемлемой. Наконец, существует убеждение, что массовые вымирания последних 50 тыс. лет вызваны охотничьим нажимом первобытного человека. Так были истреблены гигантские нелетающие птицы моа Новой Зеландии 980 ± 40–900 ± 45 лет назад [421; 480, с. 222], предположительно мамонтовая приледниковая фауна Старого Света и вся мегафауна Нового Света на рубеже плейстоцена – голоцена 11700 календарных лет назад [480, с. 221–222; 735], мегафауна Австралии 50 ± 5 тыс. лет назад в результате вторжения людей 55 ± 5 тыс. лет назад [570]. Доказано, однако, что мегафауна Австралии вымерла ок. 46,4 (51,2–39,8) тыс. лет назад из-за крайней аридности (засушливости), вызванной гляциальным максимумом [647], т. е. холодным стадиалом Вюрм II C (46,5–39 тыс. лет назад), когда вместе с похолоданием резко обострилась сухость земного климата, поскольку масса воды, в том числе атмосферной влаги, была скована в обширных ледниках. Кроме того, обнаружено, что Новый Свет был заселен ранее 33 тыс. лет назад, о чем свидетельствуют остатки первобытной культуры в Монте Верде (Чили, 33370 ± 530 лет назад) и Тока ду Бокейрану да Педра Фурада (Бразилия, св. 32 тыс. лет назад) [385; 500, с. 708], в силу чего заселение Америки не может быть увязано с вымиранием ее мегафауны 11 700 лет назад. Указанные обстоятельства не позволяют принять антропогенную гипотезу массовых вымираний, т. е. древних массовых вымираний, вызванных человеком (за вычетом новозеландского эпизода с птицами моа, который, впрочем, не относится к числу примеров массовых вымираний). Следует добавить, что вулканическая, космическая и антропогенная гипотезы массовых вымираний полностью оторваны от закономерностей развития земной жизни и не проливают здесь никакого света. Вдобавок людям, в том числе и ученым, вообще свойственно переоценивать значение катастроф в истории природы и общества и хвататься за них, как за соломинку, когда что-либо не сразу поддается объяснению (к сказанному добавим неокатастрофическое объяснение происхождения самосознания Дж. Джейнсом, см. разд. 3.2). Между тем, реальное значение катастроф не так фатально. Например, катастрофическое извержение вулкана Кракатау (Индонезия, 1883 г., погибло 36 тыс. человек) или удар Тунгусского метеорита (Красноярский край, Россия, 1908 г., произошел вывал леса в радиусе 15–30 км вокруг эпицентра) не оказали на развитие биосферы никакого влияния. В изложенной нами версии эволюции биосферы под влиянием эколого-климатического воздействия суперконтинентального цикла бросается в глаза то, что его крупномасштабная периодичность не объясняет более дробных колебаний климата. В реальности суперконтинентальный цикл создает лишь общие условия для развития оледенений или межледниковий. Их конкретные проявления подчиняются влиянию частных факторов астрономического, атмосферного, гидросферного характера, хронометраж которых пока не разработан [70, с. 20; 560; 635; 742; 750], если не считать климатических последствий от колебаний в орбитальном движении Земли с периодами в 25, 44, 100, 133 и 400 тыс. лет [602]. Хронометраж заметных вымираний приводит к выводу, что их пики отстоят друг от друга на 20–60 млн лет [184]: на 26 млн лет за последние 150 млн лет [239, с. 50; 361, с. 363; 410] или на 27 млн лет за последние 250 млн лет (8–12 пиков) [634]. Комментарии этих наблюдений пока преждевременны. Если принять нашу версию направленной эволюции, то встает вопрос, как она согласуется с дарвиновской моделью случайного хода эволюции и каков генетический механизм направленной эволюции. Мы отвечаем на него следующим образом. Земная биосфера испытывала климато-экологическое давление среды на протяжении более чем 3,85 млрд лет, и это давление как таковое являлось одним из внешних условий, к которым живые существа приспосабливались в результате случайного естественного отбора. Следовательно, их геномы давно должны были «обзавестись» наследственным механизмом ожидания климато-экологических последствий суперконтинентального цикла, т. е. фактически усвоить склонность к эволюционной направленности естественным путем. В сущности, мы остаемся в рамках дарвинизма, хотя это учение избегало представлений об эволюционной направленности. Геномы организмов слепы: они лишены органов чувств. Между тем, уже в течение нескольких миллиардов лет незрячие геномы с успехом противостоят изменчивой среде, все больше совершенствуются и, похоже, умеют предвидеть изменения в природе. Как же им это удается? Ответ здесь может быть один. Слепые геномы ориентируются в окружающем мире на основе принципов теории игр. Разумеется, крохотные геномы не сильны в теории. Однако практическая жизнь и безжалостный естественный отбор сохранили в их рядах только те образцы, которые действуют так, словно и впрямь знакомы с Дж. фон Нейманом (1903–1957, основоположником теории игр). Для разъяснения их «талантов» нам не понадобится высшая математика. Допустим, вы втянуты в азартную игру, а глаза у вас завязаны. Как вы построите свое поведение? По-видимому, пойдете с крупной карты. А если не получится, то с мелкой. Если не выйдет, ограничитесь чем-то средним. И будете так экспериментировать, пока не зацепитесь за удачу. Геномы ведут себя точно так же. Они находятся в состоянии постоянного мутагенеза, т. е. подвергают свои гены тем или иным мутациям, происходящим с примерно одинаковыми скоростями (ср., однако, разд. 2.4). Вопреки бытующему убеждению, результаты этого процесса далеко не произвольны, не случайны. Дело в том, что генные мутации делятся на серии характерных типов: гиперморфные (они увеличивают проявление подчиненного себе признака, фена), гипоморфные (они уменьшают проявление признака), антиморфные (признак уничтожается вообще) и неоморфные (признак дополняется) [7, с. 42–49]. Иными словами, мутагенез носит количественный характер (как и ваше «слепое» поведение в мысленном эксперименте с азартной игрой). Изменения блоков генов и целых хромосом также подчиняются количественному принципу: они дуплицируются, т. е. удваиваются, умножаются, как и отдельные гены [82; 468; 522; 523; 613]. Что такой мутагенез может дать геному и его организму-хозяину? Уверенность в завтрашнем дне. Допустим, грядет похолодание. Организмы слепо, механически мутируют обрисованным количественным способом. Треть из них немотивированно мутирует гипоморфно и утрачивает, например, запасы жира – они замерзнут. Другая треть вообще воздерживается от мутаций и прозябает. Зато оставшаяся треть, не подозревая о своей гениальности, опять же слепо, механически предается гиперморфным мутациям и ни с того ни с сего наращивает подкожный слой жира. Приходит холод, эта треть не мерзнет и овладевает миром. Она побеждает автоматически, в силу законов теории игр. На Земле уцелели лишь те организмы, которые неукоснительно следовали описанному способу количественного мутагенеза. Он позволял слепо предугадывать климато-экологические последствия суперконтинентального цикла и гарантировал жизнь определенной части участников событий. Другая часть так же гарантированно «не попадала» в ход событий и беспомощно вымирала. При таком положении дел в эволюции мы могли бы ожидать, что определенный процент организмов в истории жизни будет вымирать систематически и внешне беспричинно – в силу того, что эти организмы не угадали правильный ход в мутагенезе. Но так ли это? Это так. Палеонтологи действительно обнаружили в ископаемой летописи загадочную череду автоматически текущих вымираний, которую они окрестили Законом Постоянного Вымирания, или гипотезой Красной Королевы (персонажа из «Алисы в стране чудес» Л. Кэрролла) [182]. В свете вышеизложенного мы начинаем понимать, откуда в палеонтологической летописи взялся след Красной Королевы. Теперь посмотрим, что дает версия количественного мутагенеза для понимания узловых моментов истории жизни. Можно было бы ждать, что организмы, угадавшие грядущие перемены в экосреде, окажутся приспособленными к ней наиболее точно и займут в ее ландшафте господствующие высоты. Такие приспособленные организмы называются специализированными. Их судьба незавидна. Углубившись в «подгонку» себя к господствующей среде, холодной (как зверозубые пресмыкающиеся) или теплой (как динозавры), они сожгут за собой эволюционные мосты и с переменой вектора развития среды (с похолодания на потепление или наоборот) окажутся беспомощными и вымрут (как угасли зверозубые рептилии и динозавры). «Отыграть эволюцию назад» им не позволит закон Долло о необратимости эволюции, а эволюция необратима потому, что не является прямолинейной, и угадать в ней извилистые пути отступления в полном соответствии с путями наступления – нереально. Например, мы помним, что с похолоданием в ранней юре двуногие прозауроподы встали на четвереньки и сделались могучими зауроподами. Но переделать свою прожорливую мезозойскую природу они не смогли и с необратимым похолоданием в конце мела сошли с эволюционной сцены. То есть они сумели отыграть эволюцию назад лишь отчасти и поплатились за это местом под солнцем. Напротив, организмы, воздержавшиеся от углубленного приспособления к наличной среде, не пережившие подходящих количественных мутаций, прозябают в ее условиях на экологических задворках. Такими были жалкие четвероногие млекопитающие на фоне яркого мезозоя. Подобных живых существ называют генерализованными, и их участь, вопреки первому впечатлению, завидна. Когда среда меняется, им не приходится искать длинных путей попятной эволюции, поскольку они не проходили пространных путей поступательного развития. В результате они относительно легко приспосабливаются к новым условиям, довольно быстро становятся новыми господами планеты и из разряда генерализованных организмов перекочевывают в пул специализированных существ. Такое превращение произошло, как мы показали выше, с тусклыми млекопитающими на протяжении прохладного палеоцена, в начале которого канули в Лету последние динозавры. Сенсационная судьба этих колоссов объясняется их адаптацией. Вообще массовые вымирания не более невероятны, нежели мелкие [165]. Но вымирание динозавровой фауны закономерно перекрыло все допуски. Дело в том, что преобладающие организмы мезозоя были приспособлены к весьма высокой биопродуктивности среды, и ее падение в конце мелового периода смело господствующие династии животных подчистую, наводя ученых на мысли о вулканической или космической катастрофе. Напротив, звероподобные пресмыкающиеся времен пермо-карбонового оледенения ориентировались на скромный уровень биопродуктивности среды. Ее подъем в триасовом периоде сделал адаптацию зверообразных пресмыкающихся невыгодной, но не лишил их привычно обильной пищи, как это произошло с динозаврами в финале их бытия. Поэтому зверообразные пресмыкающиеся частью вымерли, частью ушли в эволюционную тень без обвального вымирания. Это обстоятельство уберегло наших предков млекопитающих от кризиса в теплом мезозое. Они даже усовершенствовались по части повышения мобильности. Можно спросить, куда же дальше, если они и без того были четвероногими? Ответ на этот вопрос таков. Одной из форм повышения мобильности животных является живорождение, поскольку оно не привязывает животных к месту кладки яиц. Во время потепления средней юры четвероногие динозавры претерпели кризис и освободили некоторые экониши, в которые устремились сумчатые млекопитающие. Они были примитивно живородящими, а потому более подвижными по сравнению с собратьями по классу. То же самое произошло с более совершенными по живорождению плацентарными млекопитающими, что в нынешнее кайнозойское время вывело их на первые биологические роли. Таким образом, можно убедиться, что наша версия стихийно направленной эволюции не состоит в противоречии с данными палеонтологии и эволюционной биологии. Достоинством этой версии мы считаем последовательное подчинение узловых эволюционных событий единообразной причинности, однотипным принципам развития. Нам кажется весьма существенным, что в описанной системе представлений предки людей – ископаемые гоминины – закономерно находят свое место, что проливает свет на стартовые условия антропогенеза. 2.4. Гоминизация Судя по молекулярно-генетическим данным, приматы вместе с ближайшими древесными родичами, предками лесных планеристов шерстокрылов [438], отделились от прочих плацентарных млекопитающих в начале кампана (верхний мел), ок. 75 млн лет назад [809, с. 1090, табл. 1]. Под влиянием похолодания в середине эпохи палеоцена, ок. 60 млн лет назад, из состава приматов выщепились мобильные, летающие формы – крыланы, или летучие лисицы [363; 619], а древолазающие формы распались на полуобезьян (тупайи, долгопяты, лемуры) и антропоидов (обезьян) [670, 112, табл. 6–7]. Последние с раннего эоцена (55–50,5 млн лет назад) обнаруживаются в палеонтологической летописи Северной Африки [178; 543; 544; 709] и Юго-Восточной Азии [236; 245; 436]. На рубеже эоцена и олигоцена от узконосых обезьян Старого Света отошли широконосые обезьяны Нового Света, 36 ± 3 млн лет назад. Наконец, в начале миоцена, 22 ± 2 млн лет назад, узконосые обезьяны разделились на низших, церкопитекоидных (павианы, макаки и т. п.), и человекоподобных, гоминоидов (гиббоны, орангутаны, гориллы, шимпанзе, люди). Вопреки расхожему убеждению, обезьяны не являются лидерами эволюционного процесса среди млекопитающих. Они – жертвы так называемого биогенетического закона, согласно которому темпы индивидуального развития животного (онтогенеза) задают скорость эволюции (филогенеза) всего вида, к которому принадлежит это животное. Причина подобной зависимости очевидна. Если индивидуальная биография особей замедлена, поколения их вида сменяются тоже медленно, и их становится меньше, чем у более активно развивающихся животных. А единицей эволюционного развития является именно поколение. Чем поколений меньше, тем медленнее протекает эволюция вида. Из-за жизни на деревьях, где нелегко воспитывать многочисленное потомство, обезьяны были ограничены в количестве одновременно рождаемых детенышей [12, с. 50–51]. Избавленные от конкуренции между собой, малыши отличались замедленным индивидуальным развитием (онтогенезом), что в силу биогенетического закона тормозило филогенез приматов, обусловливая их эволюционную отсталость. Ее признаки – цветное зрение, как у земноводных и рептилий, обильные кожные железы людей, как у земноводных, а также встречающаяся у некоторых людей шестипалость, воспроизводящая аналогичную особенность самых первых земноводных (см. разд. 2.3). У других млекопитающих подобные анцестральные, предковые черты отсутствуют. Речь, разумеется, не идет о том, что приматы вообще и в особенности люди прямиком унаследовали и сохранили перечисленные особенности амфибий. Суть дела состоит в другом. Рождая малое количество детенышей, древесные приматы затормозились в эволюционном развитии, в том числе в молекулярно-генетическом, в эволюции генов, хромосом, геномов. У людей в силу неотении эта эволюционная заторможенность усугубилась (см. ниже). В результате наш геном сохранил застойный облик – стоит его «толкнуть» и вызвать у него так называемые «обратные мутации» (т. е. изменения, восстанавливающие предковый облик отдельных генов) [82, с. 69–70], как появятся гены настолько древние, «земноводного облика», что среди млекопитающих, обладающих лишь черно-белым ночным зрением, появятся существа со зрением цветного типа, т. е. приматы. У некоторых из них вдобавок обнаружатся обильные кожные железы земноводных и даже (иногда) свойственная их ранним представителям шестипалость – это мы находим у людей, которые, таким образом, оказываются еще застойнее, нежели другие приматы, что эволюционно выгодно (парадокс направленной количественной эволюции, см. разд 2.3). В силу этого эволюционного аутсайдерства приматы имели предпочтительные шансы успешно приспосабливаться к изменениям среды (в кайнозое – к потеплениям, губительным для продвинутых млекопитающих). Напомним, что, согласно современным представлениям, «примитивные наследуют Землю» (слоган основоположника эволюционной генетики С. Оно [82, с. 174]), что обещало нашим обезьяньим предкам нелегкую, но перспективную судьбу. Указанные диалектические обстоятельства заведомо делали эволюцию приматов запутанной. В результате ни среди африканских гоминоидов (человекоподобных обезьян) [144, с. 641; 344; 350; 790], ни среди eвроазиатских дриопитеков (их родни) [144, с. 642, 643; 383, с. 69–70] не найдено надежных предков гоминин. Это привело к афро-евроазиатско-африканской гипотезе происхождения искомых предков человека [726], которая в настоящее время видится так. С началом ранне-среднемиоценового потепления, 22–13 млн лет назад, предки современных гоминоидов в предвкушении открывающихся на севере экониш (под влиянием тепла) мигрировали из Африки в Евразию, что вызвало расхождение церкопитекоидов (мартышек) и гоминоидов (предков людей). С началом похолодания среднего миоцена – раннего верхнего миоцена, 13–10 млн лет назад, исконные африканские гоминоиды (проконсулы, афропитеки, кениапитеки и т. п.) вымерли, а сородичи двуногого перелоапитека (Испания, 13 млн лет назад) вернулись из Евразии в Африку, где положили начало гоминидам (гориллам, шимпанзе, предкам людей и людям согласно систематике [383, с. 66–70], в которой люди отнесены к биологическому отряду приматов, семейству гоминид, трибе гоминин). Состав двуногих предков человека и их родственников, гоминин [131, с. 526, рис. 1; 231, с. 850, рис. 1; 144, с. 644; 383, с. 70; 821; 822], приводим ниже. Обратим внимание на то, что молекулярно-генетические даты для гоминин не всегда согласуются с палеоантропологическими датами. Они приводятся под литерой «М». Перерассчитанные с «неотеническим коэффициентом» 1,894 (мотивацию см. ниже), они даны с литерой «Н». 1. Pereloapithecus cataluniensis («обезьяна из Перелоа каталонская»). Перелоа, Каталония, Испания, 13 млн лет назад. 2. Нгорора, Баринго, Кения, 11,94–9,82 млн лет назад (ближе к нижнему пределу), 1 верхний моляр (коренной зуб) гоминида [191; 494, с. 174; 500, с. 109]. Расхождение человека с большими африканскими человекообразными обезьянами: 6,25 ± 5,75 млн лет назад (М) [143; 196; 289; 403; 445; 476, с. 25; 506; 517; 646; 670; 726; 756; 801; 809, с. 1092], или 11,84 млн лет назад (Н). 3. Sahelanthropus tchadensis («сахелец чадский»). Торос-Меналла, пустыня Джураб, северный Чад, 7–6 млн лет назад [220; 221; 775]; cp. [814]. Строение основания черепа позволяет предполагать у «сахельца» прямохождение [839]. 4. Orrorin tugenensis («первобытный человек тугенский»). Лукейно, Туген Хиллс, запад Баринго, Кения, 6,2 ± 0,19–5,625 ± 0,5 млн лет назад [494, с. 174; 623; 687]. Табарин, Туген Хиллс, запад Баринго, Кения, 4,96 ± 0,03 млн лет назад [416] (?). 5. Ardipithecus ramidus kadabba («базисная обезьяна-корень патриарх»). Аса Кома, средний Аваш, Афар, Эфиопия, 5,77 ± 0,08–5,54 млн лет назад [393; 811]. 6. Ardipithecus ramidus («базисная обезьяна-корень»). Ас Дума, Гона, Афар, Эфиопия, 4,51–4,32 млн лет назад [686]. Арамис, средний Аваш, Афар, Эфиопия, 4,387 ± 0,031–4,29 млн лет назад [797; 798; 812; 813]; cp. [145, с. 556; 687] (предок шимпанзе?). 7. Australopithecus africanus («южная обезьяна африканская»). Штеркфонтейн, Йоханнесбург, Трансвааль, ЮАР, 4,17 ± 0,14(0,35) – ок. 2,0 млн лет назад [607]. 8. Australopithecus anamensis («южная обезьяна озерная»). Канапои и Аллия Бэй, Туркана, Кения, 4,17/4,12–3,89 ± 0,02 млн лет назад [492]. 9. Australopithecus afarensis («южная обезьяна афарская»). Танзания. Эфиопия: Белодели и Када Хадар, 3,89 ± 0,02 – ок. 3,0 млн лет назад [467; 799; 36; 443]. 10. Australopithecus bahrelghazali («южная обезьяна с реки газелей»). Бахр эль Газаль, Коро Торо, Борку-Эннеди-Тибести, Чад, ок. 3,5–3,0 млн лет назад [219]. 11. Australopithecus garhi («южная обезьяна-сюрприз»). Хата, Боури, западный Аваш, Афар, Эфиопия, ок. 2,5 млн лет назад [155; 408]. 12. Australopithecus habilis («южная обезьяна умелая»). Олдувай, Танзания, ок. 1,83 млн лет назад [442; 495, с. 32–41; 708; 720; 816]. 13. Paranthropus aethiopicus («околочеловек эфиопский»). Западный берег озера Туркана, Кения, 2,53 ± 0,05 – ок. 2,45 млн лет назад [787; 283]. 14. Parantropus boisei («околочеловек бойсов»). Танзания, Кения, Эфиопия, ок. 2,5–1,3 млн лет назад [284; 286; 556; 741]. 15. Paranthropus robustus («околочеловек массивный»). Сварткранс и Кромдраай, ЮАР, ок. 1,8–1,5 млн лет назад [284; 556; 720; 737; 738]: имел способность к точному захвату, позволяющему искусно владеть орудиями? cp. [599; 758]. 16. Kenyanthropus platyops («кениец плосколицый»). Обособление линии, ведущей к роду Homo, – 1,86 млн лет назад (М) по Х-хромосоме [403], или 3,52 млн лет назад (Н). Ломекви, западный берег озера Туркана, Кения, ок. 3,5–3,2 млн лет назад [493]. 17. Kenyanthropus rudolfensis («кениец с озера Рудольфа», акселерированный австралопитецин). Чемерон и Кооби Фора, Кения, 2,43 ± 0,02 – менее 1,82 ± 0,04, 1,6 ± 0,05 млн лет назад; Олдувай, Танзания, 1,839 ± 0,005–1,785 млн лет назад; Чивондо, Малави, ок. 2,4 млн лет назад; Хадар, Эфиопия, типичный олдовай, ок. 2,3 млн лет назад [193; 417; 487; 493; 495, с. 32–41; 510, с. 419; 679; 818; 819]. 18. Homo ergaster («человек-мастер», акселерированный ранний питекантроп). В Африке 1,9/1,8–1,43 ± 0,02/1,41 ± 0,02 млн лет назад [153, с. 732, табл. 1, 733, рис. 1; 216; 217; 281; 487; 708; 741, с. 490]. В Азии: Лонггупо, Китай, олдовай, 1,942–1,785 млн лет назад [272; 426; 823]; Моджокерто и Сангиран, Ява, Индонезия, 1,81 ± 0,04, 1,66 ± 0,04 млн лет назад [507; 744]; Дманиси, Грузия, олдовай, Homo georgicus («грузинский человек»), ок. 1,75 млн лет назад [337; 338; 770]. 19. Homo erectus («человек прямоходящий», неотеничный питекантроп). В Европе: Шандалья I, Хорватия, ок. 1,6 млн лет назад [500, с. 943]; д’Орсе и Куэва Виктория, Испания, ок. 1,5 млн лет назад [352]. В Азии: Триниль, Ява, Индонезия, 0,71 млн лет назад [500, с. 1031, 1071; 487]. 20. Homo heidelbergensis («гейдельбергский человек»). В Африке: Олдувай, Танзания; Данакиль, Афар, Эритрея; Даканихило, Боури, средний Аваш, Афар, Эфиопия, ранний ашель, ок. 1,0 млн лет назад; Бодо д’Ар, средний Аваш, Афар, Эфиопия, развитой олдовай, 0,64 ± 0,03 млн лет назад; Брокен Хилл, Кабве, Замбия, поздний ашель (санго), Homo rhodesiensis («родезийский человек»), 0,11 млн лет назад [126; 154; 249; 500, с. 138, 159]. Исход из Африки: 0,63 ± 0,21 млн лет назад (М) [749], или 1,19 млн лет назад (Н). В Европе: Гран Долина, Сьерра де Атапуэрка, Испания, Homo antecessor («человек-предшественник»), 0,783 млн лет назад [188; 229]; прогрессивный неандерталец, 0,126 млн лет назад [44]. В Азии: Мугарет-эль-Зуттие, Вади Амуд, Кинерет, Израиль, финальный ашель (ябруд), Рисс III, 0,148 млн лет назад [500, с. 1148]; Нармада, Индия, поздний ашель, не бол. 0,15 млн лет назад [454]; Джинню-шань, Китай, 0,3–0,21, 0,281, 0,237, 0,187 млн лет назад; Джяньде, Китай, 0,117–0,09 млн лет назад [214]; [755]; Нгандонг и Самбунгмакан, центральная Ява, Индонезия, Homo soloensis («человек с реки Соло»), 53,3 ± 4–27 ± 2 тыс. лет назад [746]; пещера Лианг Буа, Рутенг, провинция Манггараи, о. Флорес, Малые Зондские о-ва, Индонезия, Homo floresiensis («человек с острова Флорес»), 95/74–12 тыс. лет назад [218; 584 а; 585; 572], где Homo heidelbergensis понимается по-афроевропейски узко, а потому «флоресиец» трактуется как поздний, карликовый Homo erectus, хотя лоб у малыша (рост женщины – 106 см) вздут не меньше, чем у иных «гейдельбержцев», а обезьянья челюсть не перекрывает мауэрской классики (знаменитой челюсти гейдельбергского человека из Мауэра в Германии, ок. 700 тыс. лет назад [273]) (рис. 8). 21. Homo neanderthalensis («неандертальский человек», акселерированный поздний гейдельбержец). Европа: 125 тыс. лет назад [43; 190; 285; 731]; Виндия, Хорватия, 29 080 ± 400 лет назад [128, с. 30, табл. 1]. На Ближнем Востоке: Кебара 12, Израиль, 59,9 ± 3,5 тыс. лет назад [765]; cp. [733]; Шанидар, Курдистан, Ирак, 60–44 тыс. лет назад [500, с. 968]. Расхождение с Homo sapiens – 620 ± 70 тыс. лет назад (М) [476, с. 25], или 1,17 млн лет назад (Н); ср. [614а, с. 184]. 22. Homo sapiens praesapiens («человек разумный праразумный»). Грот де Фонтешевад, Орждей, Шаранта, Франция, эвано, интер-Рисс, 281–219 тыс. лет назад (фрагмент лобной кости Фонтешевад I неотличим от соответствующей кости черепа современной европейской женщины) [500, с. 397; 480, с. 219; 767, с. 352]. Homo sapiens Omo I, Omo II («человек разумный с реки Омо», 2 образца). Формация Кибиш, южная Эфиопия, 195 ± 5 тыс. лет назад [559]. Homo sapiens idaltu («человек разумный старший»). Херто, Боури, средний Аваш, Афар, Эфиопия, ашель – средний каменный век, 160 ± 2–154 ± 7 тыс. лет назад [248; 796]; cp. [316; 732, с. 693]. Homo sapiens modernus («современный человек разумный»). По Y-хромосоме – 153,5 ± 116,5 тыс. лет назад (М) [157, с. 1364; 295; 335; 396; 793; 801; 820]. По митохондриальной ДНК – 200 ± 80 тыс. лет назад (М) [228, с. 31; 774; 801]. По ДНК – 502,5 ± 386,5 тыс. лет назад (М) [157, с. 1364; 335; 646; 781]. Среднее по темпам мутагенеза – 200 ± 80 тыс. лет назад (М), или 378,81 тыс. лет назад (Н). Расогенез по селективно нейтральной дерматоглифике (пальцевым узорам и ладонным линиям) [111], что согласно [202; 626]; cp. [782]: Негроиды – 169,6 ± 10,5 тыс. лет назад (Н); ср. 117,5 ± 82,5 тыс. лет назад (М) [157, с. 1364; 228, с. 35; 403; 451; 612; 749; 756; 820]. Класиес Ривер Маут, ЮАР, 125–105 тыс. лет назад [349; 729]; Бордер Кейв, ЮАР, св. 105 тыс. лет назад [349], 80–70, 65–50 тыс. лет назад [386], 74 ± 5 тыс. лет назад [387]. Европеоиды – 130,1 ± 4,3 тыс. лет назад (Н). Ближний Восток: Кафзех, Израиль (непрерывное обитание 20–30 тыс. лет), 90 тыс. лет назад [349], 115 ± 15, 92 ± 5 тыс. лет назад [734; 766]; Эс Схул, Израиль, 101 ± 12, 81 ± 15 тыс. лет назад [734]. Европа: пещера Бачо Киро 11, Дряново, Габрово, Болгария, ориняк I (ольшев) типа Ишталошко (нижний слой), Вюрм II Мурсхофд, 50 + 9/ – 4 тыс. лет назад (1?) [579]; Бокштайн-Тёрле, Германия, ориньяк, 46380 + 1360/ – 1170 лет назад [256]; Эль Кастильо, Испания, ориньяк, 40 ± 2 тыс. лет назад; Л’Арбреда, Испания, ориньяк, 39,9 ± 1 тыс. лет назад [728]; грот де Фе, Шательперрон, Франция, ориньяк, 39780 ± 390, 35540 ± 280 тыс. лет назад [366а]; Пештера ку Оасе, Румыния, ок. 35 тыс. лет назад [803 a, с. 332]. Калибровка радиоуглеродного возраста по методу NotCal04 дает следующие даты распространения ориньякского сапиенса: Бокер Таштит, Израиль, св. 49 тыс. лет назад; Бачо Киро, Болгария, 46 тыс. лет назад; Богунице, Чехия, 46 тыс. лет назад; Виллендорф, Австрия, 45,5 тыс. лет назад; и т. д. до Шательперрона, Франция, 41 тыс. лет назад [567 а].  Рис. 8. Человек с острова Флорес (Homo floresiensis) обитал на одном из Малых Зондских островов в Индонезии 74–12 тыс. лет назад, когда всей планетой уже владел биологический вид Homo sapiens. «Флоресиец» представлял собой позднюю карликовую форму гейдельбергского человека и достигал всего 106 см роста (судя по женскому экземпляру). Он не являлся ни сказочным хоббитом («чертенком»), ни позднейшим питекантропом, а сделался крохой подобно другим карликовым островным млекопитающим (таким, как карликовые мамонты о-ва Врангеля или вымершие карликовые слоны о-ва Мальта) [23а] Австралоиды – монголоиды – 83,9 ± 3,1 тыс. лет назад (Н). Азия: Люджянг, Китай, 67 тыс. лет назад [214]; Япония, 70–50 тыс. лет назад [160]. Австралазия: Ниах грейт кейв, Саравак, Калимантан, Малайзия, св. 41500 ± 100 лет назад; Хуон, Папуа – Новая Гвинея, 53–45 тыс. лет назад [382]; Матенкупкум, Новая Ирландия, северная Меланезия, 32700 ± 1550 лет назад [135, с. 709]; Малакунанджа II, западный откос плато Арнемленд, Австралия, 61 ± 13–45 ± 9, 52 ± 11 тыс. лет назад [648]. Америка: Монте Верде, Чили, 33370 ± 530 лет назад; Тока ду Бокейрану да Педра Фурада, Бразилия, св. 32 тыс. лет назад [385; 500, с. 708]. Представители рода Homo отличались строением тела, приспособленным к прямохождению. Они имели относительно короткие руки и относительно длинные ноги, как у двуногих динозавров. В отличие от прочих млекопитающих, наши предки обладали не слишком различающимися резцами, клыками и коренными зубами, т. е. были равнозубыми, как динозавры, что объясняется их ориентацией на высокобиопродуктивную среду, когда не требуется тщательно утилизировать пищу. Сходство усугублялось тем, что некоторые динозавры (ранние карнозавры и целурозавры) располагали хватательными верхними конечностями с отстоящим внутренним пальцем, аналогом нашего большого [25, с. 45–46, 49]. Однако, несмотря на подходящую физическую организацию, мы все еще не обладаем твердыми наследственными навыками прямохождения. Это доказывается судьбами homo ferus (детьми, выращенными животными), которые без человеческого воспитания к прямохождению не способны (ползают на четвереньках) [76, с. 47–49, 183–186; 576]. Следует добавить, что, хотя люди преодолевают стометровку не быстрее крохотной, но четвероногой крысы (ок. 10 с в обоих случаях), они все же кое в чем продвинулись как выносливые бегуны. Предки рода Homo были приспособлены к выносливому бегу гораздо хуже [207], что отвечает большей неотеничности. Способность к выносливому бегу появилась лишь у Kenyanthropus rudolfensis или Homo ergaster в результате акселерации (противоположной неотении, см. ниже). Человек современного вида перенес до трех акселераций, а потому оказался способен к марафону, хотя все еще десятикратно уступает в этом четвероногим гиенам или волкам. Представители остальных родов гоминин, будучи тоже прямоходящими, сохраняли обезьянье строение тела. Они имели относительно длинные, питекоидные руки (Australopithecus habilis, Australopithecus afarensis, Ardipithecus ramidus [36; 145, с. 556; 442; 443]), а также обезьянье строение вестибулярного аппарата, существенного для прямохождения (Australopithecus habilis, Paranthropus robustus [720]). Иными словами, история прямохождения у гоминин, от самых ранних до современных, постепенна и не закончена до сих пор. Прямохождение возникло именно у наших обезьяноподобных предков благодаря эволюционной отсталости приматов, делающей их податливыми для фундаментальных перемен. Среди млекопитающих эволюционно продвинуты не они, а мышевидные грызуны [82, с. 97]. Энергетически экономная двуногость явилась откликом на подъем биопродуктивности среды в связи с потеплением, когда подвижный поиск пищи стал неактуальным, раз она обильна и скучена в пространстве. Подобная кормовая база спровоцировала у гоминин «мутацию прожорливости». Они стали потреблять примерно в 3,8 раза больше пищевой энергии на единицу массы тела, нежели все остальные высшие млекопитающие. У последних соответствующий показатель (постоянная Рубнера) составляет 191 600 ккал на кг массы тела в течение жизни, тогда как у людей это 725 800 ккал на кг телесной массы [6, с. 73, 89; 661; 662]. Таким образом, метаболизм (обмен веществ) у людей по сравнению с другими высшими млекопитающими завышен в 3,8 раза. Неотения и акселерация Вынужденные перерабатывать почти вчетверо больше энергии, чем равновеликие млекопитающие, гоминины стали пропорционально дольше жить [323] и, соответственно, формироваться. Их индивидуальное развитие затормозилось, растянулось во времени и привело к сохранению инфантильных (детских) и даже эмбриональных черт во взрослом состоянии, что называется неотенией (от греч. ?????? – юность) (рис. 9) [10; 12, с. 17–19; 402; 577; 584, с. 13–44]. Аналогичное явление (педоморфоз) известно у земноводного аксолотля [564]. У гоминин неотения выразилась в удержании эмбриональной обезволошенности тела и невыраженности лицевого скелета, в силу чего мозговой скелет, купол черепа, непропорционально вздулся. Это не связано с интеллектуальными нагрузками: у испытывающего их последние две с половиной тыс. лет сапиенса мозг ничуть не изменился. Известно, что вздутие купола черепа началось у Australopithecus afarensis до того, как усложнилась кортикальная организация его мозга (организация его мозговой коры) [315]. Проще сказать, у гоминин обезьяний по сути мозг механически разросся в силу неотении, не усложняясь как орган интеллекта. Отметим, что старший современник «афарца», Australopithecus africanus из Таунга в ЮАР, по стадиям прорезывания зубов напоминал не ребенка 5–7 лет (как следовало бы из его вероятного возраста), а детеныша человекообразной обезьяны 3–4 лет [257, с. 626] в силу того, что фенотипическая (телесная) неотения отставала у гоминин от генотипической. Аналогичным образом наша генетическая предрасположенность к прямохождению еще не гарантирует, что человеческий ребенок усвоит двуногость автоматически. 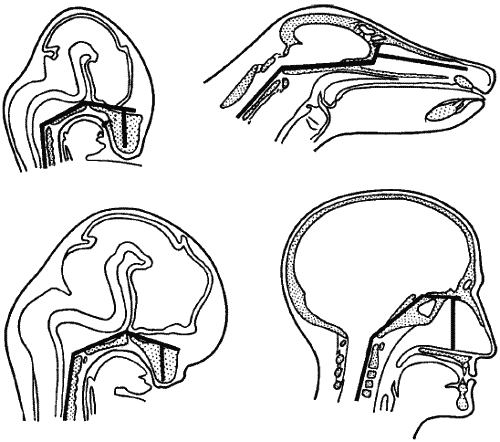 Рис. 9. Человек и его предки гоминины возникли в результате эволюции путем неотении, когда организм во взрослом состоянии сохраняет детские и даже эмбриональные (зародышевые) особенности. У человека к ним относятся обезволошенность тела (эмбрионы человека и многих других млекопитающих безволосы), а также вздутый купол черепа. У эмбрионов человека и прочих высших млекопитающих он вздут, однако после рождения у животных сильно разрастается морда, и мозговой участок черепа выглядит убогим; у людей этого не происходит: лицо остается крохотным, а потому мозг кажется обширным. По общему (ошибочному) убеждению это объясняется большим интеллектом людей. Однако человеческий ум существенно развивался только у Homo sapiens, да и то лишь последние 2,5 тыс. лет. При этом мозг человека современного типа не изменился ни на йоту. Вдобавок наш мозг активно загружен не более чем на 10%. Здесь мы видим эмбриональные и зрелые формы черепа у собаки и у человека. Оба эмбриональных абриса похожи, но лишь у человека зародышевые очертания сохраняются во взрослом состоянии [12, с. 19] Под влиянием неотении, замедлившей индивидуальное развитие наших предков, у женщин и самок гоминин месячный цикл растянулся, так что эпизоды способности к зачатию как бы слились, в результате чего эструс (аналог течки у самок животных) размазался и стерся, хотя у женщин сохраняется несколько ежемесячных дней, когда они повышенно восприимчивы к ухаживаниям. Самки гоминин стали существенно дольше, чем равновеликие млекопитающие, вынашивать потомство, что вызывало накопление ими заметных жировых запасов, необходимых для питания плода [107; 402; 583; 628]. Начиная с питекантропа, изменились сроки прорезывания зубов у молоди наших предков [280]. На протяжении своей эволюции гоминины все заметнее уподоблялись эмбрионам, что выразилось у них в грацилизации, т. е. в утончении костей скелета и приобретении изящества. Ныне это явление усиливается благодаря цивилизованному образу жизни: он вызывает последствия, напоминающие эффекты одомашнивания животных, которое изнеживает, инфантилизирует наших питомцев [490, с. 357–358]. Неотеническое увеличение продолжительности жизни у гоминин имело эволюционные последствия. Темпы эволюции зависят от скорости смены поколений. Поэтому замедление онтогенеза влечет за собой торможение филогенеза вида. У гоминин он в 3,8 раза замедлен против равновеликих млекопитающих. Поэтому, сравнивая их молекулярную эволюцию с нашей, мы должны умножать хронометраж на своего рода «неотенический коэффициент» 1,894, чтобы привести к среднему знаменателю нормально текущую эволюцию с нашей, вчетверо заниженной. Как можно видеть из вышеприведенного хронометрированного списка гоминин, подобная коррекция молекулярно-генетических дат (М) при помощи «неотенического коэффициента» дает исправленные датировки (Н), совместимые с палеоантропологическими данными. Заторможенность молекулярного мутагенеза у человека как таковая выявлена эмпирически [506], однако она не дотягивает до заторможенности человеческого онтогенеза, что объясняется сложностью расчетов абсолютных количественных показателей для мира млекопитающих, где они зависят от размеров, физиологии и других особенностей животных. Так, продолжительности жизни кошки, человека и кита относятся примерно как 1:10:20, в то время как массы их тел относятся друг к другу как 1:10:1500, откуда видно, что рост продолжительности жизни у млекопитающих заметно отстает от увеличения их размеров. Принятое нами отношение древности современного человека (378,81 тыс. лет) к возрасту гоминин (11,84 млн лет) составляет 1/31,25, в то время как ожидаемое достигает почти 1/25 [100, с. 12]. Данное противоречие объясняется тем, что современный человек перенес три акселерации против двух у своих предшественников (см. далее). Это должно было ускорить его молекулярную эволюцию в 1,5 раза – вышло 1,25, ибо последняя, текущая акселерация только началась. Проще говоря, в истории наших предков имелись эпизоды ускоренной эволюции (акселерации), отчего их молекулярно-генетический возраст оказался ниже ожидаемого (т. е. возраста ок. 474 тыс. лет). В сравнении с армадой двуногих мезозойских динозавров прямоходящие гоминины выглядят сиротливо. Двуногие прыгающие сумчатые (например кенгуру), насекомоядные (например прыгунчики) или грызуны (например тушканчики) прибегают к энергоемкой скачущей локомоции. Она быстра, но не экономична. Выгодной ходящей или бегающей локомоцией ныне обладают лишь бегающие птицы. Она выгодна, но не проворна. Так, африканский страус при массе тела 70–90 кг и соответствующей мощи развивает скорость всего 70 км/ч, тогда как гепард с массой тела 50–65 кг способен на спринт до 112 км/ч. Четвероногие ходоки и бегуны легко покрывают огромные пространства (например волки 500–600 км за день), что благоприятно в нынешних условиях скромной, разреженной пищи. В мезозое ее плотность была в среднем выше, а потому отбор дал плеяду прямоходящих динозавров. Сейчас он поощряет четвероногих животных, а гомининам отводит роль исключения. Отсюда наша аномально высокая постоянная Рубнера и нестандартная эколого-эволюционная судьба. Живя дольше равновеликих млекопитающих, гоминины накапливали в своих сообществах представителей разных поколений и демографически умножались, что продолжается по сей день. Организмы в биомах подчиняются популяционным волнам, когда при благоприятных условиях последовательно умножаются сперва растения, потом растительноядные потребители, а затем хищники. Далее растительноядные истощают кормовую базу, падают в числе и вызывают упадок хищников. Раз за разом процесс повторяется [80, с. 243–258; 97, с. 62–72]. Поэтому поголовье животных в биомах в среднем фиксировано. Медленно развиваясь и размножаясь, гоминины не успевали за популяционными волнами, а потому постепенно, но неуклонно росли в численности. Их демографическая история объясняет ряд новаций в эволюционном состоянии и образе жизни. Напомним, что для видообразования требуется репродуктивная (по воспроизводству себе подобных) изоляция меньшинства от большинства [82, с. 93]. Напротив, высокие плотности населения затрудняют эволюционные изменения, отчего многочисленное современное человечество не эволюционирует [82, с. 96]. Прямохождение, высокий метаболизм и неотения возникли у наших предков в условиях низкой плотности населения, а затем фенотипически прогрессировали до наших дней. Условием для такого порядка вещей были умеренные плотности населения. Между тем в силу неотении гоминины были склонны к повышению своей численности. При ее пиковых изменениях, демографических взрывах следовало бы ожидать процессов, обратных неотении. Неотения – это замедление развития. Поэтому при демографических всплесках приходилось бы ожидать явлений ускорения развития, т. е. акселерации. Демография Однако гоминины не могли выдерживать длительных периодов своей повышенной численности, поскольку их перенаселение истощало кормовую базу, подрывало равновесие со средой и угрожало им крахом, ведь они были эволюционно заторможены и не могли быстро изменяться, восстанавливая свою численность. Выжили те их сообщества, которые стихийно научились стабилизировать свою населенность. К концу оледенения Бибер I (3,2–3,0 млн лет назад), чреватого для Африки аридностью (засушливостью и скудостью), гоминины должны были попасть в депрессию, а их сообщества изолироваться, что благоприятствовало видообразованию. В плювиальный (влажный и обильный) интерстадиал Бибер I/II (3,0–2,6 млн лет назад) гоминины размножились в новом видовом составе, так что на хронологической точке в 2,5 млн лет назад они представлены новыми видами Australopithecus garhi, Paranthropus aethiopicus и, возможно, Kenyanthropus rudolfensis. Хотя последний известен лишь с отметки 2,43 ± 0,02 млн лет назад, свойственная ему археологическая культура типичного олдовая началась еще св. 2,63 ± 0,5, 2,58 ± 0,23 млн лет назад. Если предположить, что гоминины человеческой линии (т. е. Kenyanthropus) ощутили вышеуказанные эколого-демографические трудности, то естественным способом обойти их мог стать для «кенийцев» уход в другую экологическую нишу. Биомасса растений в биоме примерно в 10 раз превосходит биомассу растительноядных потребителей, а биомасса последних – в 5 раз биомассу хищников [80, с. 86–87]. Подобно близкородственным шимпанзе (в особенности шимпанзе-пигмею бонобо [667]), древние гоминины были всеядными существами с преобладанием растительной диеты [139; 277]. Если бы они обратились к хищничеству, их численность автоматически упала бы в 5 раз (условно), что нейтрализовало бы их неотеническое размножение, в 3,8 раза превышающее свойственное равновеликим млекопитающим. Мнение о том, что гоминины обратились к некрофагии (трупоядению) [13; 18, с. 180–195; 90, с. 107–113; 711]; cp. [501; 772], неприемлемо, поскольку экониша падальщиков в силу своей вторичности теснее экониши хищников, и такая конкуренция была бы не по силам преимущественно растительноядным гомининам, мало пригодным к борьбе за падаль, например, со львом (50%-ный падальщик) или гиеной (30%-ный падальщик). Проще сказать, активным охотникам доводится бороться лишь с добычей, которая не всегда способна постоять за себя, в то время как падальщикам постоянно приходится сражаться с другими некрофагами, среди которых имеются смертельно опасные фигуранты (по степени опасности в Африке это львы, гиены и леопарды, которые совмещают потребление падали с умелой активной охотой, а потому являются грозными соперниками). Кроме того, в эконише некрофагов попросту меньше пищи, нежели в эконише охотников за живой добычей. Вдобавок даже состоявшуюся падаль стремительно пытаются растащить и некрофаги скромных габаритов (шакалы, грифы, стервятники). Шимпанзе (а также павианы) около 3% пищи добывают групповой охотой [18, с. 183; 59, с. 142–143, 147; 103, с. 180, 182, табл. 6.8; 194; 722]. Трудно сомневаться, что гоминины, известные впоследствии как великие охотники, на заре своей истории совпадали по этой части с шимпанзе. «Кенийцы», усугубив свое пристрастие к животной пище, угодили в эконишу хищников и сократились в численности. Возможно, их путем пошли и прочие гоминины, например Australopithecus africanus (любил листву и/или сочные фрукты [380], а также мясо [719]), отличавшийся от земляка Paranthropus robustus (всеяден [349], с пристрастием к грубой пище [380]). Решение демографической проблемы посредством перехода в эконишу хищников оказалось временным, поскольку «кенийцы» продолжали размножаться и испытывать прежние эколого-демографические трудности, еще более опасные для хищников, нежели для травоядных (хищники малочисленнее и уязвимей). В разное время эта незадача привела к вымиранию прочих австралопитецинов. «Кенийцы» же развили питекоидные способности к производительному применению орудий, впервые в истории животных сделали их коллективными и положили начало демографо-технологической зависимости, связывающей плотность населения со сложностью технологии (см. разд. 3.1). Осваивая орудия археологической культуры типичного олдовая (2,6 млн лет назад) и переживая неблагоприятный гляциал Бибер II (2,6–2,3 млн лет назад), «кенийцы» некоторое время пребывали в стабильном состоянии. Однако с наступлением мягкого интергляциала Бибер/Донау (2,3–2,0 млн лет назад) они вновь размножились, открыв первый в истории человечества демографический взрыв. Около 2,2 млн лет назад он выплеснул «кенийцев» из Африки в Евразию, где их типично олдовайские орудия отмечены в Диринге (Якутск, Якутия, 3,2/3,1–2,7/2,2 млн лет назад [75, с. 131]), в Сен-Валье (Дром, Франция, 2,2 млн лет назад [500, с. 935]) и в Сент-Эбле (Мон Купе, Франция, 2,5–2,2 млн лет назад [127, с. 28]). Высокая плотность населения вызвала в среде «кенийцев» акселерацию и увеличение размеров тела, отчего Kenyanthropus rudolfensis долгое время воспринимался в палеоантропологии как крупный вариант Australopithecus habilis [815]. После этого эпизода «кенийцы» до конца ледниковой стадии Донау I (2,0–1,9 млн лет назад) около 300 тыс. лет снова пребывали в эволюционном покое, поскольку разрядили демографическую обстановку предпринятой экспансией. Умеренная плотность населения вернула им полноту неотении, очередным этапом которой стало появление нового гоминина с обостренной неотеничностью. Это выразилось в увеличении его мозгового черепа, укорочении верхних конечностей (инфантильный признак) и усовершенствовании вестибулярного аппарата, обслуживающего прямохождение, а также в сапиентных темпах взросления, судя по стадиям прорезывания зубов. Иначе говоря, возник питекантроп. Он сразу же испытал рост плотности населения, пережил второй демографический взрыв и 1,835 млн лет назад мигрировал в Евразию, где известен от Индонезии до Испании (исходим из того, что размеренное расселение от Восточной Африки до Индонезии занимало 25 тыс. лет [507], а в Индонезии «человек-мастер» появился 1,81 млн лет назад). Прохождение им Левантского коридора отмечено олдовайской индустрией в Эрк-эль-Ахмар (долина Иордана, Израиль, 1,95–1,77 млн лет назад) [656]. Под влиянием повышенной плотности населения он испытал вторую акселерацию и предстал как Homo ergaster, по существу классический питекантроп акселерированного типа. Его африканский представитель, образец Нариоко-томе III (Туркана, Кения, 1,60 ± 0,05 млн лет назад [216; 217; 708]), имея возраст 12 ± 1 год, уже достигал роста 1,68 м, что обещало в зрелости ок. 2 м. Его азиатский представитель из Сангирана (Ява, 1,66 ± 0,04 млн лет назад) тоже известен как гигант, «мегантроп». Наконец, ребенок из Моджокерто (Ява, 1,81 ± 0,04 млн лет назад) в 1 ± 0,5 года имел объем черепа 72–84% от взрослого питекантропа. Такое соотношение юных и зрелых особей присуще шимпанзе и на 15% опережает развитие человеческого ребенка [261], т. е. является акселерированным. Кстати, акселерированный Homo ergaster до 2 м роста – типа Нариокотоме III или «мегантропа» – представляется наиболее вероятным кандидатом на роль гипотетического «снежного человека», скрытно распространенного в горах Центральной Азии и лесах Северной Калифорнии. Прямых данных о росте плотности населения у африканского Homo ergaster нет. Погибшее при наводнении сообщество Australopithecus afarensis в Хадаре (Эфиопия, 3,5–3,0 млн лет назад) насчитывало ок. 13 индивидов [36, с. 158, 225]. Носители олдовайской культуры в Олдувае (Танзания, ок. 1,8 млн лет назад) жили сообществами в 15 ± 5 индивидов. Спустя миллион лет этот показатель удвоился, и сообщество Homo heidelbergensis в Олоргесаилие (Кения, 0,74–0,60 млн лет назад) насчитывало 30 ± 20 индивидов [50, с. 87–88; 249]. Поскольку данных о демографических взрывах у «гейдельбержцев» нет (см. далее), можно думать, что часть этого удвоения населения приходилась на долю демографического всплеска у «человека-мастера», а другая часть обусловила медленное расселение «гейдельбержца» в Евразию ок. 0,783 млн лет назад. Подчиняясь демографо-технологической зависимости, Homo ergaster в силу возросшей плотности населения породил ашельскую технологию (1,6–1,5 млн лет назад), после чего пришел в демографически-стабильное состояние, вернулся к неотеничному облику и предстал как классический Homo erectus, менее рослый, но более головастый. Углубление этого инфантильного облика переродило его в Homo heidelbergensis, известного в Африке ок. 1,0 млн лет назад. Несколько ранее, в интерстадиале Гюнц I/II (1,27–0,93 млн лет назад) 1,19–1,17 млн лет назад (Н) на стадии позднего питекантропа или раннего гейдельбержца, разошлись пути предков сапиенсов и неандертальцев. В теплую межледниковую стадию Гюнц/Миндель II (790–770 тыс. лет назад) ок. 783 тыс. лет назад Homo heidelbergensis достиг Европы, где предстал в Испании как Homo antecessor. Около 300 тыс. лет назад он появился в Китае, освоил Индонезию и закончил свой путь на Яве как Homo soloensis уже в эпоху азиатских сапиенсов. Поскольку равномерное расселение покрывает расстояние от Восточной Африки до Явы всего за 25 тыс. лет [507], передвижения гейдельбержца нельзя назвать демографическим взрывом. Сопутствующих примет акселерации он также не обнаруживал. В теплую межледниковую эпоху Рисс/Вюрм (144–110 тыс. лет назад) финальный гейдельбергский человек в Европе, «прогрессивный неандерталец», достиг таких плотностей населения, что принял акселерированную форму Homo neanderthalensis (классический неандерталец), обозначившуюся 125 тыс. лет назад. Этот гоминин был на 30% крупнее нынешнего среднего человека и даже на 24% превосходил массивных обитателей наших высоких широт [449, с. 125], что отражало ускорение онтогенеза у неандертальца [541]. Наши высокоширотные современники акселерированы в силу того, что у млекопитающих в суровых условиях поднимаются скорости основного метаболизма (обмена веществ) [291], в результате чего организм развивается быстрее (этот повседневный метаболизм не следует смешивать с пожизненным метаболизмом, способным растягивать биографию организма, см. выше.) Неандертальский человек был столь форсирован в развитии, что приобретал питекоидные черты черепа еще на пренатальной стадии, в утробе матери [627]. Его зубные коронки формировались на 15% быстрее, чем у современных людей, а зрелость наступала в 15 лет [658, с. 937]. По темпам акселерации он не уступал Homo ergaster из Моджокерто. О демографическом взрыве и территориальных конфликтах у неандертальцев свидетельствует каннибализм (cp. [163]). Холодный интерстадиал Вюрм II B 1–2 (59–51тыс. лет назад) толкнул разросшуюся популяцию неандертальцев к миграции на юг, где 59 тыс. лет назад они соприкоснулись с ближневосточными сапиенсами. Подчиняясь демографо-технологической зависимости, европейские неандертальцы в эпоху Вюрм II/III (37,5–36 тыс. лет назад) создали прогрессивную верхнепалеолитическую культуру перигор 0, сменившуюся перигором I (шательперроном), обозначенным в эпохи Вюрм II/III и Вюрм III А 1 (37,5–32,5 тыс. лет назад). В конце холодного стадиала Вюрм III B (30–29 тыс. лет назад) неандертальцы, окруженные процветающими сапиенсами, прекратили свое существование. Популяция Homo heidelbergensis из Бодо д’Ар в Эфиопии (640 ± 30 тыс. лет назад) в начале интергляциала Миндель/Рисс (388–333 тыс. лет назад) настолько углубилась в неотению, что породила так называемую популяцию «Евы», 378,81 тыс. лет назад (Н), представители которой отличались особо вздутым куполом черепа и, соответственно, относительно убористым лицом, что является характерными анатомическими признаками вида Homo sapiens. Он представлял собой, таким образом, этап инфантилизации гейдельбергского человека (в противоположность неандертальцу, который являлся акселерированной стадией гейдельбергского человека). В теплую интерстадиальную эпоху интер-Рисс (281–219 тыс. лет назад) ранние сапиенсы (пресапиенсы) посетили Европу, где оставили каменные орудия прогрессивной культуры эвано верхнепалеолитического облика. Их представители типа Фонтешевад I не снискали будущего в Европе. В эпоху значительного оледенения Рисс III (219–144 тыс. лет назад) развившаяся пустыня Сахара отделила североафриканскую часть популяции «Евы» от субсахарской. Последняя положила начало негроидной расе современного человека, 170 тыс. лет назад (Н), представленной автралоидным Homo sapiens idaltu (160 ± 2–154 ± 7 тыс. лет назад) [732, с. 693] и монголоидообразными людьми койсанской (бушменской) расы из Класиес Ривер Маут и Бордер Кейв в ЮАР, 125–105 тыс. лет назад. Австрало-монголоидный облик, вероятно, был типичен для популяции «Евы», что проявилось в конце ее существования (см. ниже). В теплом Рисс/Вюрме (144–110 тыс. лет назад) она двинулась на север и, положив начало европеоидной расе на Ближнем Востоке 130 тыс. лет назад (Н), отбыла на восток. Ее отпрыски в Израиле представлены протокроманьонцами из Кафзех (115 ± 15–92 ± 5 тыс. лет назад) и Эс Схул (101 ± 12–81 ± 15 тыс. лет назад). Достигнув Индии, популяция «Евы» распалась на австралоидов и монголоидов, разделивших ее анатомические черты 84 тыс. лет назал (Н). Свидетельством ее пребывания на субконтиненте является полиморфизм (разнообразие) щелочной фосфотазы плаценты и других признаков у местного населения, поскольку он характерен для центра распространения биологического вида [92], который в случае с человеческими расами переместился из Восточной Африки в Индию. Как отмечалось выше, популяция «Евы» сочетала австралоидные и монголоидные признаки, которые обусловили облик Homo sapiens idaltu и представителей койсанской расы. Они же предрешили типажи австралоидной и монголоидной рас после расщепления популяции «Евы» в Индии. Можно допустить, что она отличалась и европеоидным началом, которое на различных стадиях распада протопопуляции сообщило сходные черты европеоидам и америндам (американским индейцам), в частности развитое строение носа, хотя у европеоидов оно основано на костной, а у америндов – на хрящевой ткани [2, с. 158; 3, с. 27]. Тем временем у европеоидов на Ближнем Востоке произошел демографический взрыв, и они вторглись в Европу, используя потепление интерстадиала Вюрм II Мурсхофд (51–46,5 тыс. лет назад). В силу демографо-технологической зависимости всплеск их народонаселения породил пластинчатую технологию ориньякской археологической культуры (50 тыс. лет назад). Освоив Европу, пребывающие на пике демографического подъема сапиенсы вытеснили неандертальцев из выгодных охотничьих угодий и положили конец их виду (29 тыс. лет назад). Причина эволюционного успеха сапиенсов и поражения неандертальцев коренится в том, что у первых демографический взрыв состоялся позже (ок. 50 тыс. лет назад), нежели у вторых (ок. 125 тыс. лет назад), и перекрыл результаты их размножения. В силу того же демографического подъема европеоиды вошли в фазу акселерации и дали популяцию абри де Кро-Маньон (Лез Эйзиде-Тейяк, Дордонь, Франция, ориньяк IV, 27 680 ± 270 лет назад [567, с. 463]). Эти кроманьонцы в узком смысле слова отличались средним ростом 185 см у мужчин, в чем не уступали современным акселератам. Следующий, преднеолитический, демографический взрыв случился с европеоидами тоже на Ближнем Востоке 16 тыс. календарных лет назад с окончанием первой местной мезолитической культуры кебары (21,8–16 тыс. календарных, или 19–13,95 тыс. радиоуглеродных лет назад). В это время в Леванте (Израиль, Иордания, Ливан, Сирия) начался распад общности носителей шумерского, сино-кавказских и ностратических языков, т. е. почти всех известных языков белой расы, а также некоторых групп восточных и центральноафриканских негроидов, северных и дальневосточных монголоидов, северо-западных америндов [52, с. 65–73]. В соответствии с демографо-технологической зависимостью этот популяционный взрыв вызвал неолитическую технологическую революцию 11,7 тыс. лет назад. Однако признаки акселерации у тогдашних левантийцев не известны. Они ожидались бы у носителей третьей мезолитической культуры региона, натуфийской (14,3–11,7 тыс. календарных, или 12,45–10,2 тыс. радиоуглеродных лет назад), созданной первыми афразийцами (предками древних египтян, евреев, арабов и пр.). Между тем, средний рост натуфийцев-мужчин не превышал 1,60–1,71 м [762, с. 50], что исключает акселерацию. Возможно, она проявилась в неолитическое или более позднее время, однако данных на этот счет нет. Можно лишь отметить, что исторические западные семиты из Передней Азии отличались большей рослостью, нежели, например, древние египтяне. Наконец, последний демографический взрыв произошел в Западной Европе исторически недавно. Его первый этап относится к XI – середине XVI вв. [23, с. 67–75]. Умножение западноевропейского населения ознаменовалось крестовыми походами на Восток (1096–1270 гг.), германской экспансией и Великими географическими открытиями кон. XV – XVII вв. Благодаря демографо-технологической зависимости вслед за демографическими событиями последовала промышленная революция, открывшаяся мануфактурами XIV в. в Италии. С кон. XVIII в. в Западной Европе развертывается акселерация роста и созревания населения [12, с. 423–427]. Во 2-й пол. XX в. эта тенденция отчетливо охватила Северную Америку, Австралию, Россию и Восточную Азию, т. е. приобрела глобальные масштабы. Данное биологическое явление представляется нормальной эволюционной реакцией человечества на рывок демографического подъема. Отметим, что плавное демографическое развитие Индии, Юго-Восточной Азии и Китая никогда не сопровождалось процессами акселерации и действием демографо-технологической зависимости. Исключение составляет эпоха тамошней неолитической революции (в технологическом плане), а также современная акселерация в Китае и Японии (см. разд. 3.1). Красота Неотения отразилась не только на биологической и технологической истории гоминин. Она затронула врожденные человеческие чувства прекрасного, связанные с оценкой физического облика себе подобных, т. е. с оценкой их физической красоты [53, с. 13–15]. Общеизвестно и подкреплено специальными исследованиями [616], что у людей физически привлекательны признаки здоровья, плодовитости и властности. Эти особенности привлекательны для особей противоположного пола у всех позвоночных и многих беспозвоночных животных. Следовательно, названные особенности не могут считаться специфически человеческими, и мы не станем на них останавливаться. Мы попытаемся раскрыть природу физической красоты, присущей лишь человеку и, вероятно, его гомининным предкам. Вопреки существующему убеждению, природа человеческой красоты не является тайной. Она обусловлена биологически и воспринимается инстинктивно. Как мы видели, в своей истории гоминины неоднократно проходили неотенические и акселерированные стадии развития. Неотеничность отвечает эволюционной направленности гоминин. Поэтому она репродуктивно (сексуально) привлекательна. Однако чередование неотеничных и акселерированных эпизодов привело к тому, что представления о красоте раздвоились. Красота лица неотенична, поскольку эволюционно перспективна. Красота тела акселерирована, поскольку акселерация создает механические трудности при деторождении. Проще говоря, в нормальной человеческой популяции присутствуют как низкорослые неотеничные индивиды, так и высокорослые акселераты. Они свободно скрещиваются. Поэтому некрупная неотеничная женщина может родить ребенка от акселерата. Однако женский таз – это жесткая конструкция. Он затрудняет производство акселерированного потомства. Поэтому женщины независимо от роста имеют большой объем таза. У акселераток он органичен. У неотеничек – вынужден. В любом случае биологически выгодно быть первыми. Поэтому они инстинктивно оцениваются как гармоничные, красивые. Отсюда интуитивно (по привлекательности) выстраиваются кроманьонские параметры современных топ-моделей. Это высокорослые женщины ок. 180 см высоты, с объемами 90–60–90, что отражает характерные параметры акселерации – как нынешней, так и древней, кроманьонской. Высокорослые мужчины красивы в силу соответствия подобным женщинам. Мы не станем говорить о банальной выгодности высокорослости при охоте и т. п. – хотя бы потому, что все зависит от того, где и на кого охотиться. Как известно, в джунглях охотнику выгоднее быть пигмеем. Происхождение притягательных черт лица гораздо проще. В результате неотении череп наших предков стремился во взрослом состоянии сохранять эмбриональную модель, т. е. быть по возможности сферичным. Для решения этой задачи лицевой отдел черепа ушел под лоб, надбровья выпрямились и наехали на глазницы, скулы, нос, челюсти и зубная дуга сократились. В результате сформировалась женская классическая или кукольная внешность. Ее образцами отличаются, например, Ким Уайлд (английская певица с весьма выразительным лицом) или Магда Конопка (западная актриса польского происхождения, типологически близкая Ким Уайлд и вдобавок носившая титул «леди Вселенная»). Мужская красота в своей основе та же, только массивнее в силу полового диморфизма (полового расхождения). Образцами красивой мужской внешности обладали, например, лорд Байрон (великий английский поэт) или Рудольф Валентино (знаменитый актер немого кино). Можно разобрать черты лица в деталях. Красив высокий закругленный лоб – потому, что его форма оптимально вписывается в сферичную неотеничную модель. Все прочее представляется угловатым и негармоничным. Красивы вытянутые глаза в низких глазницах – потому, что в древности округлые обезьяньи глазницы наших предков в результате неотении сплюснулись у гоминин под наехавшим лбом, отчего стали низкими и визуально продолговатыми. Отсюда же притягательность прямых бровей, бровей вразлет и т. п. форм. Оптимален небольшой прямой или чуть вздернутый нос (у женщин), находящийся подо лбом. Он хорошо вписывается в сферичную, неотеничную модель. Миниатюрные, утопленные в лицо скулы очаровательны по той же причине, как и облегченные челюсти, подбородок и зубная дуга. Понятно, у мужчин все должно выглядеть рельефнее, «мужественнее». Очерченные критерии физической красоты, в принципе, однотипны во всех расах. Однако следует помнить их специфику. Негроиды оформились в резулультате вздутия черепного свода (общего для всех рас), миниатюризации скул и носа. Челюсти отставали от процесса, и это называется прогнатизмом (сильное выступание тяжелых челюстей вперед). С европеоидами случилось то же, лишь уменьшились челюсти (ортогнатность, утопленность челюстей под скулы), а нос остался велик. Монголоиды отличаются плоскими, но широкими скулами, а австралоиды сохраняют общую грубость лица, впрочем усредненно сократившегося как целое. Неотеничные критерии красоты следует применять к расам с указанными поправками. Непоследовательность в формировании лиц по расам объясняется тем, что все они независимо произошли от исходной популяции «Евы», перемещавшейся из Восточной Африки в Индию. Как можно видеть, магистральные пути гоминизации у наших предков укладывались в поступательную неотению или отклонения от нее (Homo ergaster, Homo neanderthalensis). Неотения, прямохождение и высокая постоянная Рубнера (а также связанная с ней прожорливость) одинаково вытекали из эволюционной направленности гоминин в сторону экономной маломобильности, что проявляется в склонности к гиподинамии и ожирению у современного человека. Если генетическое обеспечение гоминизации состоялось на заре трибы гоминин, то фенотипическое воплощение заложенных в генетической программе тем не завершилось до сих пор. Это выражается в ряде примет количественной эволюции (см. разд. 2.3). У человечества продолжаются редукция (измельчание) зубов и сокращение зубной дуги, равно как и общее облегчение всего скелета (грацилизация). Углубление внешних проявлений неотении ведет ко все большей миниатюризации человеческого лица, отчего грубые лица среднему человеку инстинктивно кажутся несовременными, «пещерными», что в определенном смысле отвечает сути дела. Наконец, развитые (и не только) страны страдают от гиподинамии и ожирения населения, что объясняется развертыванием заложенной в нас генетической программы. Гиподинамия представляется опасным, но адекватным воплощением нашей эволюционной маломобильности, а ожирение – досадным выражением одного из последствий неотении (напомним, что ожирение у женщин обусловлено аномально растянутой беременностью; у мужчин же избыточный вес прибывает в силу генетической солидарности с прекрасным полом, обусловленной общим для обоих полов генотипом). Последняя пара примеров эволюционно чрезмерна, невыгодна. Однако в гигантском человечестве естественный отбор подавлен, а потому на свет беспрепятственно выходят такие наши генетические задатки, которые в небольшой, активно эволюционирующей популяции давно бы пресеклись. Понимание природы гоминизации имеет мировоззренческое значение. Человек стремился и стремится познать самого себя, как призывал еще Хилон Лакедемонский, третий из семи греческих мудрецов [104, с. 92–93]. Осуществить эту мечту, наблюдая себя и современников, не представляется возможным. История технологического человечества занимает более 2,6 млн лет. За это время природные гоминины подверглись таким сложным превращениям, что истинная суть покрылась флером наивных мифов. Приведем красноречивый пример. Едва ли кто-то сомневается, что человек обязан интеллектом своему большому мозгу. При этом упускается из виду, что: homo ferus (дети, воспитанные животными), располагая большим мозгом, не имеют никакого интеллекта, человеческий мозг задействован не более чем на 10%, эпоха интеллектуального расцвета человечества (последние 2,5 тыс. лет) не сопровождалась изменениями головного мозга человека. Вывод очевиден. Размеры мозга не обусловливают интеллект. Они, скорее всего, развились у людей как побочный результат неотении. Без знания гоминизации этого вывода не сделать. Тогда откуда же у людей интеллект? Чтобы ответить на этот вопрос, нам придется уяснить, какова природа технологического прогресса, чем полезен интеллект, помимо его изобретательности (присущей и животным), и как, наконец, людям удалось создать громаду земной цивилизации. |
|
|||
|
Главная | Контакты | Прислать материал | Добавить в избранное | Сообщить об ошибке |
||||
|
|
||||