|
||||
|
Генетика и феногенетика поведения животных[39] Генетические исследования поведения имеют существенное значение для ряда областей биологии и медицины. Во-первых, они должны быть той базой, на которой только и могут развиваться некоторые области физиологии высшей нервной деятельности. Учение об индивидуальных различиях высшей нервной деятельности (в том числе учение о ее типах) и выяснение относительной роли врожденных и индивидуально приобретенных компонентов поведения невозможны без генетического анализа. Вполне понимая это, И. П. Павлов создал в Колтушах лабораторию генетики высшей нервной деятельности. Во-вторых, генетика дает возможность при помощи скрещиваний разъединять и соединять в гибридном потомстве те и другие особенности поведения с различными морфофизиологическими свойствами организма и выяснять корреляционные зависимости между теми и другими. Это открывает новый тонкий метод изучения зависимости формирования поведения от морфофизиологических свойств организма, который невозможен при помощи современного хирургического или физиологического метода. В-третьих, изучение генетики поведения имеет большое значение для ряда проблем эволюционного учения. Генетически обусловленные особенности поведения животных играют роль в структуре популяций. Наследственные различия в поведении обусловливают образование изолированных популяций различной численности, что имеет большое значение для темпа эволюционного процесса. В-четвертых, изучение генетики поведения животных важно для отыскания новых методов наиболее рационального одомашнивания хозяйственно полезных животных. Это имеет особенно большое практическое значение для пушно-звероводческих хозяйств. В-пятых, генетика поведения необходима для создания экспериментальных моделей нервных болезней. У мышей описано несколько десятков неврологических наследственных болезней, которые изучаются в качестве экспериментальных моделей болезней человека. Генотипически обусловленная модель эпилепсии широко изучается на грызунах во всех странах. В 1965 г. этому вопросу был посвящен интернациональный коллоквиум во Франции. Генетические исследования особенностей поведения начались вскоре после вторичного открытия законов Менделя. Накопившийся к настоящему времени материал показывает, что многие особенности поведения наследуются по законам Менделя, однако в большинстве случаев ряд факторов изменяет картину их наследования. Для генетических исследований поведения удобной моделью оказались оборонительные реакции у животных. Этому вопросу посвящен ряд исследований. 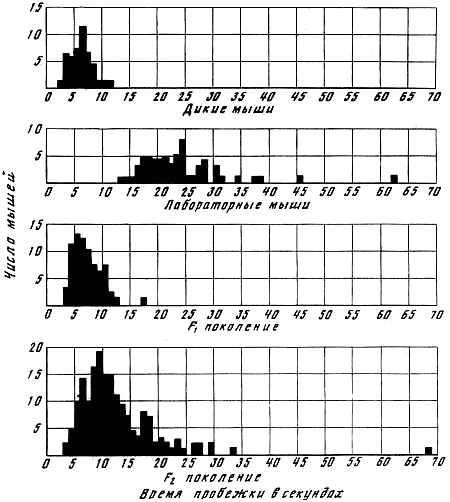 Рис. 1. Наследование пугливости у мышей Рис. 1. Наследование пугливости у мышей В 1932 г. Давсоном были проведены исследования способа наследования резко выраженной пугливости диких мышей по сравнению со слабой выраженностью этого признака у лабораторных мышей. Всего было исследовано 3376 особей. Использовался объективный метод регистрации: время пробежки по коридору (длиной 24 фута) при пугании мыши передвигающимся движком. Предварительное исследование выявило высокую корреляцию (r=+0,92+0,003) между отдельными испытаниями одних и тех же мышей, указывающую на значительную стабильность изучаемых признаков поведения. Среднее время пробежки для диких мышей составляло 5 с, для домашних — 20 с. В первом поколении наблюдалось почти полное доминирование пугливости диких мышей. Среди особей второго поколения значительно увеличилась вариабельность в степени пугливости (рис. 1) по сравнению с F1. На основании своего исследования Давсон пришел к выводу, что различие в пугливости между дикими и домашними мышами определяется двумя или тремя генами. Практически все дикие мыши гомозиготны по этим доминантным генам. Помимо основных генов, определяющих степень пугливости родительского поколения, на формирование изучаемых признаков оказывают влияние несколько модификаторов. Это исследование показало наследование особенностей поведения по законам Менделя, однако оно в то же время проиллюстрировало, что это наследование осуществляется, как и в случае большинства количественных различий между признаками, при участии полимерных генов. Наследуемость особенностей поведения по тем же законам, по каким наследуются морфологические признаки, ясно указывает, что эволюция поведения осуществляется в результате естественного (или искусственного) отбора наследственных изменений. На это указывал еще Ч. Дарвин в главе об инстинктах в «Происхождении видов». В настоящее время накопился значительный материал, подтверждающий воззрения Дарвина по этому вопросу. В качестве примера, показывающего роль селекции в изменении характера поведения, могут быть приведены работы по геотаксису у Drosophila melanogaster. На рис. 2 приведены результаты селекции по изменению геотаксиса. Селекция в течение 65 поколений привела к дивергенции: были созданы линии с ясно выраженным положительным и отрицательным геотаксисом. Обратная селекция (проводившаяся между 52-м и 64-м поколением) повела к изменению характера геотаксиса. На основании гибридологического анализа авторы приходят к выводу о полигенной природе произошедших изменений в поведении мух, которые зависят от генов, расположенных в аутосомах и Х-хромосоме. Наряду с гибридологическим анализом различий в особенностях поведения очень существен феногенетический метод, позволяющий устанавливать механизм наследственного осуществления генотипически обусловленных признаков. Примером простой зависимости наследования различных особенностей поведения от морфологических признаков является выбор температурного оптимума у мышей. Так, например, в работах Гертера было показано, что дикие мыши и альбиносы выбирают различную температуру во время отдыха. Оказалось, что температурный оптимум диких мышей — 37,36°, белых — 34,63°. Обнаружена простая картина наследования этого оптимума. Изучение показало, что температурный оптимум определяется густотой меха и толщиной эпидермиса на коже живота мыши. У белых мышей густота меха меньше, чем у диких (число волосков на единицу площади 45:70, а толщина эпидермиса больше — отношение 23:14). Особенно ясная зависимость установлена между температурным оптимумом и густотой шерсти. У гибридов Fi; температурный оптимум близок к оптимуму белых мышей: он равен 34,76±0,12°, густота шерсти — 43,71 волоска на единицу площади. 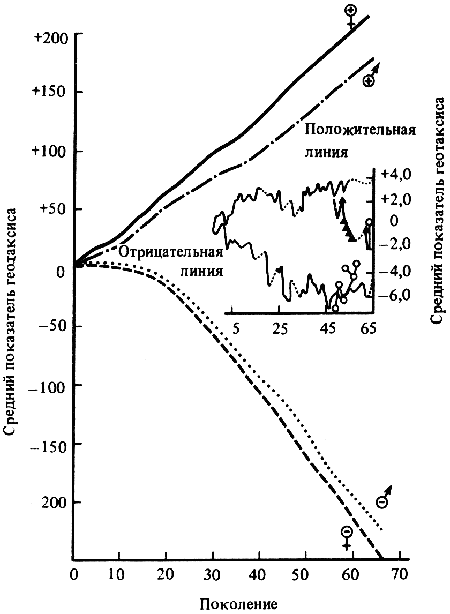 Рис. 2. Результат селекции на положительный и отрицательный геотаксис у Drosophila melanogaster (суммарные кривые) Рис. 2. Результат селекции на положительный и отрицательный геотаксис у Drosophila melanogaster (суммарные кривые) В уменьшенном масштабе приведены результаты обратной селекции (селекция мух с наиболее отрицательным геотаксисом в положительной линии и наиболее положительным в отрицательной линии). Жирные кривые — селекция на отрицательный геотаксис; тонкие — на положительный. Точками обозначены участки кривой, отображающие поколения, по которым отсутствуют данные (по Эрленмайер-Киммлингу и др., 1962). При обратном скрещивании (F1 ? дикие мыши) произошло расщепление на две группы. У одной группы оптимум соответствовал таковому у белых мышей (+34,56°±0,12) при густоте меха 52,7 волоска, в другой группе температурный оптимум был близок к оптимуму диких мышей (37°); У этой группы число волосков на единицу площади составляло 70,94. Воспитывая мышей при различной температуре, Гертер пришел к выводу, что, помимо наследственно обусловленной толщины эпидермиса и густоты шерсти, определяющих температурный оптимум, наблюдается и модификационное приспособление каждой мыши к той температуре, в которой она воспитывается. Это модификационное приспособление может изменить характерный для данной особи оптимум выбора места отдыха. Этот пример ясно показал зависимость формирования адаптационной реакции поведения от генотипически обусловленных морфологических признаков организма. Примером, в котором при помощи генетического метода удалось отдифференцировать наследование типа поведения от наследования морфологических признаков, явилась работа Мазинг по изучению фотореакции у Drosophila melanogaster. Исследование показало, что путем отбора в течение 26 поколений не удалось выделить линию, в которой бы все особи реагировали или не реагировали на свет. Это указывает на неполное проявление генов, обусловливающих разную активность мух по отношению к свету. Мухи с редуцированными глазами (Ваг, Ваг еуеless) реагируют на свет, однако реакция у них замедленная. Среди безглазых особей из линии eyeless встречаются мухи, которые оказываются активно реагирующими на свет. Это показало, что глаза не являются единственным рецептором, воспринимающим свет. Резкое ослабление фотореакции, отмеченное у мух с редуцированными крыльями, давало основание предположить, что фоторецепторы расположены на крыле. Опыты с обрезанием крыльев у мух из линии, активно реагирующей на свет, привели к значительному ослаблению положительного фототаксиса. Это подтвердило предположение о значительной роли поверхности крыльев в осуществлении фотореакции мух. Однако генетический анализ показал, что это, очевидно, не так. Система генетического анализа проводилась таким образом, что мухи vestigial скрещивались с нормальной линией мух, активно реагирующей на свет. Положительный фототропизм оказался полностью доминантным признаком. Мухи vestigial, появляющиеся в F2, скрещивались опять с нормальными мухами. После 17 поколений «вкрещиваний» гена vestigial в нормальную линию, когда генотипическая среда гена vestigial была практически заменена генотипом нормальных мух, оказалось, что мухи с редуцированными крыльями стали активно реагировать на свет. Это показало, что слабая фотореакция мух vestigial определяется нередуцированностью крыльев. Данное исследование подтвердило взгляд некоторых энтомологов, что восприятие света осуществляется всей поверхностью тела, а не связано специфически с глазами или с поверхностью крыла. Примером очевидной зависимости наследственно обусловленного поведения от различий активности половых желез могут служить исследования Мак-Гилла и Бляйта, в которых было показано, что наступление времени половой активности самцов, приводящее к спариванию после предыдущего спаривания (с эякуляцией), чрезвычайно различно в разных линиях мышей. У мышей линии C57BL/6 это время составляло в среднем 96 ч, а в линии DBA/2–1ч. Быстрое восстановление половой активности доминантно. При обратном скрещивании F1 ? C57 BL/6 время восстановления половой активности равнялось в среднем 12 ч; при этом наблюдалась большая вариация, указывающая на расщепление по этому признаку. При изучении оборонительных реакций удалось обнаружить зависимость наследственного осуществления признаков поведения от различного функционального состояния организма. Исследования, проведенные на собаках, показали, что пассивно-оборонительная реакция (пугливость, трусость), проявляющаяся по отношению к различным внешним факторам, обусловлена генотипом. В наших работах изучалось оборонительное поведение у собак по отношению к человеку. У взрослых собак при одинаковых условиях это свойство является довольно константным. Коэффициент корреляции между двумя оценками, сделанными с промежутком 1–2 года, равен +0,87±0,04. Материалом для генетического исследования послужили две группы собак: первая группа (224 особи) состояла в основном из немецких овчарок и эрдельтерьеров, воспитанных в разнообразных условиях (питомники и частные лица); вторая группа включала 89 собак, в основном беспородных. Все собаки этой группы воспитывались в условиях питомника Института физиологии им. И. П. Павлова в Колтушах. Проведенное исследование показало, что боязнь человека у собак — генотипически обусловленный признак, имеющий доминантный или неполностью доминантный характер наследования. Проявление и выражение этого свойства поведения находится в большой зависимости от ряда условий. Другая оборонительная реакция, активно-оборонительная (реакция агрессии или злобности по отношению к незнакомому человеку), исследовалась нами на 121 потомке, полученном от разных типов скрещиваний. Критерием агрессивного поведения являлось хватание зубами протянутого собаке незнакомым человеком предмета. Все собаки по этому признаку делятся на две альтернативные группы. Коэффициент корреляции, вычисленный между отдельными оценками этого признака с промежутками 1–2 года (r=+0,79±0,04), иллюстрирует довольно большую константность проявления этого признака поведения. Проведенный анализ указывает на генетическую обусловленность этой особенности поведения, имеющей доминантный характер наследования. Скотт в 1964 г. опубликовал работу по наследованию реакции лая у различных пород собак. Им установлены большие различия между отдельными породами. Наибольшие различия обнаружены между коккер-спаниелями, которые лают очень часто, и бейсенджи (африканские охотничьи собаки), которые почти не лают. Автор объясняет найденные различия разным порогом реакции собак на внешние раздражители. У спаниелей он очень низок, у бейсенджи высок. Гибриды F1 по своей реакции лая близки к спаниелям. Это указывает на доминантный характер наследования данного свойства. Характер расщепления гибридов показал, что найденные различия легче всего объяснить наличием одного доминантного гена, обусловливающего низкий порог реакции лая на внешние раздражители. Однако, помимо наследования основного гена, на формирование данного признака поведения оказывают влияние большое количество модификаторов и внешние условия. Пассивно- и активно-оборонительная реакции наследуются независимо. Если они проявляются у одной и той же особи, то формируется своеобразное злобно-трусливое поведение. При резком выражении один из этих компонентов поведения может полностью подавить проявление другого. Это было показано применением гибридологического анализа с параллельным использованием фармакологических препаратов, меняющих степень выражения отдельных компонентов оборонительного комплекса злобно-трусливых собак. Анализ наследственного осуществления оборонительных реакций поведения показал, что их проявление и выражение находятся в большой зависимости от степени общей возбудимости животного. Оказалось, что генетически обусловленные реакции поведения могут не проявиться в фенотипе животного при малой его возбудимости. Однако потомство, полученное при скрещивании таких животных с возбудимыми особями проявляет ясно выраженное оборонительное поведение. 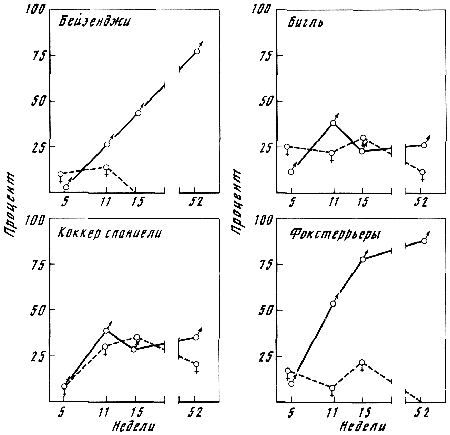 Рис. 3. Формирование с возрастом различной степени «преобладающего» поведения одного пола над другим у собак разных пород Рис. 3. Формирование с возрастом различной степени «преобладающего» поведения одного пола над другим у собак разных пород По оси абсцисс — возраст собак; по оси ординат — процент преобладающего поведения собак одного пола над другим, вычисляемого на основании времени завладения костью, даваемой двум разнополым собакам, находящимся в помещении для испытания в течение 10 мин (по Павловскому, Скотту, 1956). Повышенная возбудимость наследуется, как это было показано на разных животных, как доминантный или неполностью доминантный признак. У собак повышенная возбудимость наследуется как доминантный или неполностью доминантный признак. Повышенная двигательная активность крыс является генотипически детерминированным признаком и наследуется в качестве неполностью доминантного признака. У кур породы леггорн, по данным Головачева, порог долгосрочной возбудимости (реобаза) двигательных нервных волокон седалищного нерва превосходит таковой у кур породы австролорп и является доминантным признаком, зависящим от ограниченного числа генов. Зависимость проявления трусливого поведения у собак от разной степени возбудимости можно проиллюстрировать, на примере скрещивания немецких овчарок с гиляцкими лайками. Маловозбудимые нетрусливые гиляцкие лайки были скрещены с возбудимыми нетрусливыми немецкими овчарками. Все потомки этого скрещивания (п = 25) обладали повышенной возбудимостью и резко выраженной трусостью (рис. 3). В этом скрещивании генотипически обусловленная пассивно-оборонительная реакция была унаследована от гиляцких лаек, у которых она не проявлялась из-за недостаточно высокой возбудимости их нервной системы. Наличие у них подпороговой пассивно-оборонительной реакции было доказано искусственным повышением возбудимости их нервной системы. После введения кокаина у гиляцких лаек проявилась пассивно-оборонительная реакция. Но не только проявление, но и степень выражения наследственно обусловленной реакции поведения животных находится в зависимости от степени общей возбудимости их нервной системы. Изменяя состояние возбудимости нервной системы, можно изменять и выражение оборонительных реакций поведения. При повышении степени возбудимости в результате введения фармакологических или гормональных препаратов (гормона щитовидной железы) параллельно с увеличением возбудимости происходит и усиление оборонительного поведения. И наоборот, удаление щитовидной железы, уменьшающее степень возбудимости животного, приводит к ослаблению оборонительных реакций. Зависимость проявления и выражения генов, определяющих данные формы поведения, от генотипически детерминированной активности желез внутренней секреции была изучена нами на примере наследования пугливости диких (норвежских) крыс при скрещивании их с лабораторными крысами. При использовании в качестве теста быстроты пробежки 6-метрового коридора при пугании крысы звуковым раздражителем было установлено, что пугливость диких крыс почти полностью доминировала у гибридов F1. При обратном скрещивании (F1 ? лабораторные альбиносы) происходило ясное расщепление. Дикие (норвежские) крысы имеют по сравнению с лабораторными относительно больший вес надпочечников за счет более сильно развитого кортикального слоя. У гибридов F1 относительный вес надпочечников до 3-месячного возраста оказался промежуточным между размером надпочечников родителей. В более старшем возрасте относительный размер надпочечников приближается к размерам надпочечников лабораторных крыс. К этому возрасту уменьшается повышенная возбудимость и пугливость гибридов F1. Удаление надпочечников, а также гипофиза, приводящее к редукции надпочечников, приводило к ослаблению или даже полному устранению пугливости у полуторамесячных гибридов F1. Это ослабление пугливости происходило на фоне резкого снижения общей возбудимости. Таким образом, помимо наследования генов пугливости диких крыс, происходит, видимо, наследование повышенной функциональной активности коры надпочечников, которая обусловливает повышенную возбудимость гибридов F1. Этот пример иллюстрирует наличие некоторых генотипически обусловленных морфофизиологических взаимоотношений, которые обусловливают наследственное осуществление признаков поведения. 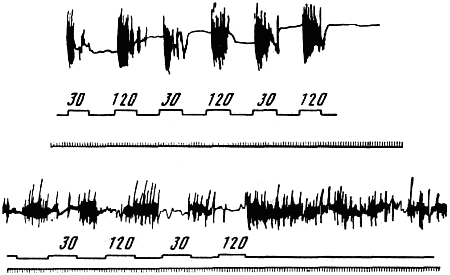 Рис. 4. Затяжное двигательное возбуждение крысы после звуковой экспозиции (нижний рисунок) и его отсутствие (верхний рисунок) Рис. 4. Затяжное двигательное возбуждение крысы после звуковой экспозиции (нижний рисунок) и его отсутствие (верхний рисунок) Нижняя кривая — отметка действия раздражителей (20 — слабый, 130 — сильный); верхняя кривая — запись двигательной активности животного (по Савинову и др., 1964) Биохимический анализ наследственного осуществления оборонительного поведения и степени общей активности у мышей проводится Маасом. В этом исследовании изучались две линии мышей: С57 BL/10 и BALB/C Мыши первой линии проявляют большую активность, меньшую пугливость и более высокую степень агрессивности по отношению к особям своего вида, чем мыши второй линии. Исследовалось содержание серотонина (5-гидрокситриптамин) и норадреналина — медиаторов возбуждения нервной системы. Исследование показало, что в стволе мозга (мост, средний и промежуточный) у линии С57 BL/10 содержится меньше серотонина, чем у BALB/C. У первых — 1,07±0,037 мг/г, у вторых — 1,34±0,046 мг/г; разница статистически достоверна: Р < 0,01). Достоверных различий в содержании норадреналина не обнаружено. Уровень содержания серотонина в определенных отделах мозга (особенно в гипоталамусе) играет роль в «эмоциональном» поведении животного. Опыты с введением фармакологических препаратов, которые меняют различные звенья обмена серотонина, показали, что найденные генетические различия в содержании серотонина у обеих линий мышей связаны с различными механизмами связывания этого нейрогормона нервной тканью. У мышей линии BALB/C происходит более быстрое освобождение серотонина нервной тканью, чем у мышей линии С57 BL/10. Эти исследования интересны в том отношении, что указывают новые пути возможной биохимической реализации генотипа в формировании особенностей поведения. Как ни велико значение гуморальных факторов и взаимоотношения отдельных физиологических систем в наследственном осуществлении признаков поведения, однако определенный тип поведения детерминирован уже на клеточном уровне. Уайтингом в 1932 г. описаны различия в половом поведении паразитических ос Habrobracon. Самки этих ос парализуют жалом личинок мельничной огневки Ephestia. Самцы не реагируют на личинок Ephestia, а ухаживают за самками Habrobracon. Такое половое различие в поведении обусловливается хромосомным набором клеток мозга. Это видно из того, что половые мозаики ведут себя по отношению к Ephestia в зависимости от генетической конституции мозга, а не в зависимости от половых желез. В том случае, если в ткани мозга осы наблюдается гинандроморфизм (участки клеток самцового типа перемешаны с участками клеток самок), то поведение таких особей резко нарушается: они пытаются ухаживать за личинками Ephestia и жалить самок Habrobracon. Такое нарушение полового поведения происходит в результате полного извращения реакции гинандроморфных особей на внешние раздражители. Чрезвычайно существен для феногенетики поведения вопрос о взаимоотношении индивидуально приобретенных и врожденных факторов. Принципиально этот вопрос не отличается от одной из основных проблем феногенетики: влияния генотипа и внешних факторов на формирование морфологических признаков. Удобным примером для рассмотрения относительной роли врожденных и индивидуально приобретенных факторов в формировании поведения являются оборонительные реакции собак. Первой работой, выполненной сотрудниками И. П. Павлова в Колтушах в 1933 г., было изучение влияния на поведение собак различных условий их воспитания. Выржиковский и Майоров, разделив два помета щенков беспородных собак на две группы, воспитали их в различных условиях. Одна группа воспитывалась в изоляции, другая — в условиях полной свободы. В результате выросшие собаки первой группы обладали резко выраженной трусостью, собаки второй группы ею не обладали. И. П. Павлов давал этому факту следующее объяснение: щенки обладают «рефлексом естественной осторожности» по отношению ко всем новым раздражителям; этот рефлекс постепенно затормаживается по мере знакомства со всем многообразием внешнего мира. Если щенок не встречается с достаточным количеством многообразных раздражителей, он остается на всю жизнь трусливым (одичалым). Последующие исследования по изучению влияния генотипа на проявление и выражение трусости в зависимости от условий содержания, проведенные нами на собаках, показали взаимодействие генотипических и внешних факторов в формировании оборонительного поведения. Материалом для этого исследования послужили немецкие овчарки и эрдельтерьеры (п = 272). Собаки обеих пород воспитывались в разных условиях: одна группа — у частных лиц, где имелась возможность соприкасаться со всем многообразием внешнего мира, другая — в питомниках, где собаки находились в значительной изоляции от внешних условий. В табл. 1 приведены данные по проявлению и выражению трусости у этих собак. При изолированном воспитании процент особей, имеющих пассивно-оборонительную реакцию, увеличивается в обеих группах. Однако среди немецких овчарок по сравнению с эрдельтерьерами значительно возрастает количество трусливых особей с резким выражением этого свойства поведения (это различие статистически достоверно). Таблица 1. Проявление и выражение пассивно-оборонительной реакции у собак разных пород, воспитанных в различных условиях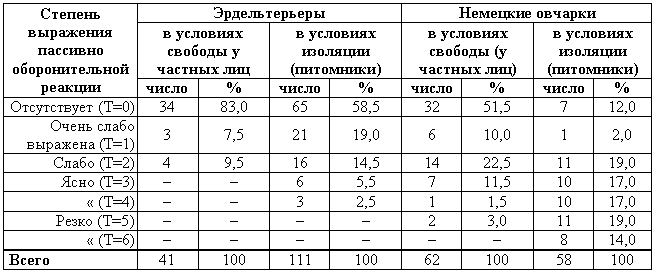 Приведенные данные свидетельствуют о том, что проявление и степень выражения трусливого поведения собак, воспитывающихся в изоляции, осуществляются в зависимости от генотипа животного. Таким образом, та «одичалость» собак, которая выражается в пугливости особей, воспитанных в изолированных условиях, является определенной нормой реакции нервной системы, обусловленной генотипом, на условия их воспитания. Несмотря на то что одомашнивание собаки началось не позднее 8000–10000 лет тому назад (Scott, Fuller, 1965), современная собака сохранила генотипически обусловленную тенденцию к одичалости, которая легко выявляется даже при незначительно изолированных условиях воспитания. Вывезенные в ненаселенную человеком территорию собаки дичают, как это произошло, например, на Галапагосских островах, куда они были доставлены испанцами для уничтожения коз, которые являлись источником пищи для английских пиратов. Эта популяция одичалых собак существует в настоящее время. Отловленные человеком щенки легко приручаются. Различие в форме выражения пассивно-оборонительной реакции свободно живущих популяций в зависимости от разного генотипа было описано Леопольдом в 1944 г. Исследовались дикие индейки, домашние и гибридная популяция. Эта популяция была получена от скрещивания диких и домашних индеек и свободно жила в штате Миссури. Дикие индейки, обнаружив на большом расстоянии опасность, сейчас же улетают. Особи гибридной популяции, обнаружив опасность, близко подпускают к себе пришельца и, отлетев на две сотни ярдов, начинают спокойно пастись. Птенцы диких индеек имеют резко выраженную тенденцию к затаиванию при приближении врага. У гибридной популяции эта форма поведения ослаблена: птенцы имеют тенденцию убегать или улетать при приближении к ним на близкое расстояние. Студнитцем описаны также различия в степени боязни человека у одних и тех же видов птиц, живущих в разных географических стациях. Наиболее ярким примером такого различия боязни человека являются дрозды-дерябы (Turdus viscivorus). В Англии они совершенно не пугливы, в Северной Европе очень пугливы, хотя и не преследуются человеком. Наличие в популяции генотипических различий, обусловливающих формирование пассивно-оборонительной реакции поведения, является, несомненно, одним из важнейших факторов быстрой перестройки оборонительного поведения при появлении врага. Такой пример был описан Ф. Нансеном. В 1876 г., когда первые суда норвежского промыслового флота проникли глубоко на Север, к берегам Гренландии, тюлени (Cystophora cristata) настолько не боялись людей, что их убивали ударом по голове. Однако по прошествии нескольких лет они стали пугливы: не всегда подпускали даже на ружейный выстрел. Подобный процесс усиления боязни человека происходит почти всегда там, где человек проникает в ранее ненаселенную территорию и начинает охоту за животными местной популяции. Конечно, помимо отбора особей с наиболее резко выраженной пассивно-оборонительной реакцией и особей, которые легче других научились бояться человека, большую роль при указанной выше перестройке оборонительного поведения в популяции может играть традиция. Все многообразные сигналы тревоги, прямое подражание поведению родителей могут явиться наряду с отбором системой ненаследственной перестройки поведения популяции. Однако, если нет наследственно обусловленных предпосылок к проявлению пассивно-оборонительной реакции, один традиционный опыт, видимо, не в состоянии обусловить боязнь врага, истребляющего данную популяцию. Такой случай описан Ю. Гексли на примере гусей Фольклендских островов. Несмотря на интенсивное истребление человеком, они проявляли лишь небольшие признаки боязни человека, не предохраняющие популяцию от истребления. Наличие генотипических различий в формировании пассивно-оборонительной реакции является предпосылкой для перестройки оборонительного поведения популяции. Установлено, что усиление пассивно-оборонительной реакции происходит не за счет прямого наследственного закрепления реакций испуга уцелевших особей, а за счет естественного отбора наиболее трусливых особей или тех, которые имели генотип, способствующий наиболее быстрому формированию пугливости при их преследовании. Без специального генотипического анализа трудно отдифференцировать наследственную и ненаследственную изменчивость поведения, традиции от поколения к поколению. Это издавна создавало условия для необоснованных допущений прямого наследования приобретенных навыков. Даже такой строгий и объективный исследователь, как И. П. Павлов, в очень осторожной форме допускал возможность наследования результатов индивидуального опыта. В 1913 г. он писал: «… можно принимать, что некоторые из условных вновь образованных рефлексов позднее наследственностью превращаются в безусловные» (С. 273). К тому же периоду относится его более определенное высказывание по этому вопросу в «Лекциях по физиологии»: «Передаются ли условные рефлексы по наследству? Точных доказательств этому нет, до этого наука еще не дошла. Но надо думать, что при длительном периоде развития прочно выработанные рефлексы могут становиться врожденными» (С. 85).[40] В начале 20-х годов И. П. Павлов поручил своему сотруднику Студенцову изучить наследование условных рефлексов у мышей. На эти опыты, не давшие положительных результатов, часто ссылались, причисляя И. П. Павлова к сторонникам наследования приобретенных признаков. Это вынудило И. П. Павлова в письме к Гуттену изложить свое отношение к этому вопросу (Правда. 1927. 13 мая). В течение всей дальнейшей жизни И. П. Павлов стоял на строго генетических позициях. Им была создана в Колтушах лаборатория по изучению генетики высшей нервной деятельности, перед зданием которой рядом с памятником Декарту и Сеченову был поставлен памятник Грегору Менделю. В качестве постоянного консультанта в своих генетических исследованиях И. П. Павлов пригласил крупнейшего генетика-невропатолога С. Н. Давиденкова, он консультировался также с Н. К. Кольцовым. Генетическая работа велась путем селекции в отдельных семействах собак по типологическим особенностям их высшей нервной деятельности. Результаты этих исследований, показавшие роль генотипа в формировании типологических свойств высшей нервной деятельности, были опубликованы после смерти Павлова. Эти исследования показали, что генотипические факторы играют значительную роль в формировании типологических особенностей высшей нервной деятельности. В различных семействах собак, в которых производилась селекция в направлении степени силы (или слабости) процесса возбуждения, наблюдалась корреляция по этим особенностям нервной деятельности между братьями и сестрами в отдельных пометах собак: r=+34±0,1. Результаты скрещиваний между собаками с сильной и слабой нервной системой, которые были начаты еще при жизни И. П. Павлова, сведены в табл. 2. Степень силы нервной системы связана с полом: самцы (п = 31) обладают более сильной нервной системой, чем самки (п = 22). Вероятность соответствия Р(х2) оказалась меньше 0,05, что может быть признано статистически значимым. Вопрос о роли генотипических факторов в обучаемости исследуется начиная с известных работ Иеркс и Багга. Иеркс изучала обучаемость двух линий крыс: одну неинбридированную, другую инбредную из института Уистара. Результаты этой работы показали, что среднее время обучаемости неинбридированных линий несколько меньше, чем инбредных (52,25 урока у первых и 65,00 у вторых). Багг исследовал индивидуальные и семейные различия в поведении мышей при нахождении пути в довольно простом лабиринте. Сравнивалась линия белых мышей, которую Багг основал в 1913 г. (линия С, в настоящее время BALB/C), с линией желтых мышей. Оказалось, что среднее время обучаемости белых мышей за 15 опытов составляло 27,5±2,0 с с 9 ошибками за урок; время обучаемости желтых мышей равнялось 83,0±7,0 с с двумя ошибками за урок. Наблюдалось сходство в обучаемости особей одних и тех же пометов. Таблица 2. Данные по наследованию сильного и слабого типа нервной деятельности у собак[41]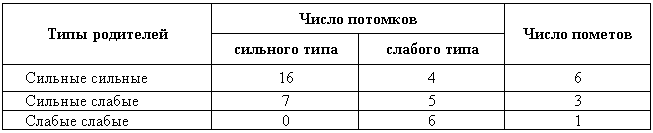 Детальное исследование роли генотипических факторов в выработке условных рефлексов было проведено Викари. В этом исследовании была изучена обучаемость японских танцующих мышей (Mus Wagneria asiatica) (инбридинг 20 лет), трех линий обычных мышей (Mus musculus), альбиносов Багга (инбридинг 14 лет), ослабленно-коричневых (инбридинг 17 лет) и ненормальноглазых (abnormal x-ray eyed) (инбридинг 6 лет). Оказалось, что каждая линия мышей имеет характерную для нее кривую обучаемости. Скрещивание между особями отдельных линий показало, что быстрая обучаемость доминирует над более медленной обучаемостью. Автор указывает, что характер расщепления во втором поколении позволяет предположить, что различие в обучаемости между коричневыми и альбиносами Багга обусловлено монофакторально, хотя нельзя отрицать и возможности более сложной картины наследования. Различия в обучаемости между линией Багга и японскими танцующими мышами обусловливаются, по мнению автора, наличием нескольких наследственных факторов. Проведенное исследование (выполненное на 900 мышах) указывает на большую роль генотипических факторов в быстроте обучаемости у мышей. Однако найденные различия при анализе такого комплексного признака, каким является быстрота обучаемости, еще не доказывают генотипически обусловленных различий в интимных механизмах мозга, связанных с самим процессом обучения. Наличие генотипически обусловленных различий в безусловнорефлекторных реакциях изучаемых линий может в значительной мере определить найденные различия в обучаемости. В качестве подобного примера можно привести исключительно тщательную работу М. П. Садовниковой-Кольцовой. Проведя изучение обучения в лабиринте (хамптонкурт) у 840 крыс, автор путем селекции вывел две линии: одну быстро обучающуюся, другую медленно. Индекс быстро обучающихся крыс (логарифм времени, затраченного на 10 счетных опытов) равен 1,657±0,025, медленно обучающихся — 2,642±0,043. Разница между обоими индексами (Д=0,985±0,05) оказалась в 20 раз больше вероятной ошибки. Дальнейший анализ показал, что найденные различия между обеими линиями крыс обусловливаются не различиями в их способности к выработке условных рефлексов, а большей пугливостью второй линии крыс (которая происходила в значительной мере от диких норвежских крыс). При обучении в аппарате Стона, в котором крыса подгонялась захлопывающимися дверцами и потому не могла из-за пугливости забиваться в угол лабиринта, обучение обеих линий шло одинаково. Таким образом, селекцией отбирались не генотипы, способствующие более или менее быстрому обучению, а генотипы, обусловливающие различную степень пугливости, которая и изменяла кривую обучаемости. Со сходным примером зависимости наследования скорости образования положительных условных рефлексов кур и осетровых рыб от безусловных рефлексов мы встречаемся в работе Пономаренко, Маршина и Лобашова. Наследование особенностей возбудительного процесса, который является одним из основных параметров при выработке условных рефлексов, авторы объясняют их корреляцией с характером безусловных рефлексов. У рыб имеется ясная зависимость скорости образования условных рефлексов от уровня возбуждения пищевого безусловного центра, зависящего, в свою очередь, от наследственно обусловленного темпа роста. Эти примеры показывают, что при генетическом анализе такого общего комплексного свойства, каким является обучаемость необходим самый тщательный и, если это возможно, параллельно проводимый физиологический анализ. В течение многих лег генетикам известно, что некоторые гены оказывают плейотропный эффект на морфологические признаки и особенности поведения. В 1915 г. Стертевант обнаружил, что рецессивный ген, расположенный на одном конце Х-хромосомы у Drosophila melanogaster и вызывающий желтую окраску тела вместо нормальной серой, уменьшает также копулятивную способность самцов. Дальнейшие исследования показали, что редуцированная половая активность самцов этой линии связана с нарушением времени и способа «ухаживания» за самками перед копуляцией. Желтые самцы, подсаженные к самкам, начинают ухаживать за ними в среднем через 9,6 мин, нормально окрашенные — через 4,9 мин. Для того чтобы началась копуляция, самцы желтой линии ухаживают в среднем в течение 10,5 мин, нормальные — в течение 6,0 мин. Помимо этого, у самцов желтой линии нарушается один из основных признаков ухаживания за самками — вибрация крыла, направленного в сторону самки. Этот акт поведения самца является необходимым ритуалом, который самка воспринимает посредством своей антенны для того, чтобы быть подготовленной к копуляции. У самцов желтой линии вибрационные удары слабее, чем у нормальных самцов, и осуществляются с более длинными интервалами. В линиях желтых мух у самок наблюдается повышенная (статистически достоверная) готовность к копуляции по сравнению с нормальными самками, являющаяся компенсаторным приспособлением для возможности нормального спаривания. Эта повышенная готовность к спариванию у желтых самок не является плейотропным эффектом гена желтой окраски. Она определяется селекцией других генов, снижающих порог копулятивной готовности. Этот пример интересен в том отношении, что показывает, как в результате одной мутации с плейотропным эффектом, меняющим реакцию поведения, и последующего отбора создается линия, которая, как считает автор этой работы Басток, может привести к возникновению физиологически изолированного экотипа. Яркий пример плейотропного действия генов на морфологические признаки и особенности поведения крыс описаны Килером и Кингом. Изучая различные мутации окрасок шерсти, проявляющиеся у диких (норвежских) крыс, содержащихся в течение многих поколений в неволе, авторы обнаружили, что мутантные особи отличаются от диких по своему поведению. Особенно резко по своему оборонительному поведению отличались мутантные особи с черной окраской шерсти. Такие крысы не кусались. Авторы считают, что полученные ими данные указывают на один из возможных путей, по которому шла доместикация диких крыс. Они предполагают, что лабораторные альбиносы возникли не в результате длительной селекции мелких мутаций, изменивших их «дикое» поведение, а в результате нескольких мутаций, некоторые из которых имели плейотропный эффект на окраску шерсти. Значительную роль в доместикации крыс сыграл отбор гена черной окраски шерсти в комбинации с геном пегости. У большинства лабораторных альбиносов эти гены находятся в криптомерном состоянии и не проявляются из-за отсутствия у альбиносов основного фактора пигментации. Исследования, выявившие наличие широкого плейотропного эффекта генов, влияющих на поведение, проводятся Беляевым и Трут на лисах. Исследования на серебристо-черных лисицах, разводимых в звероводческих хозяйствах, показали большую гетерогенность популяции по оборонительным реакциям поведения. Выделено три основных типа оборонительного поведения: активно-оборонительное (агрессивное), пассивно-оборонительное (пугливое) и спокойное (отсутствие обоих типов оборонительного поведения). Результаты поставленных скрещиваний показали, что наибольший процент лисиц с тем или другим характерным поведением наблюдается в потомстве родителей, характеризующихся таким же типом поведения: наибольший процент агрессивных лисиц рождается в потомстве от скрещивания агрессивных особей; трусливые потомки обнаруживаются в наибольшем проценте при скрещивании между собой трусливых родителей. Селекция на «спокойное» поведение оказалась эффективной. Существенно отметить, что проведенный анализ не позволяет говорить о влиянии того или другого типа оборонительного поведения матери на характер поведения потомства, которое могло сформироваться в результате подражания. Значительный процент особей в звероводческих хозяйствах составляют особи с трусливым поведением, что является вероятным результатом изолированного воспитания (клеточное содержание) лисиц. Исследование половой активности самок (время наступления течки) и их плодовитости показало, что у спокойных самок течка наступала во всех возрастных группах раньше, чем у агрессивных особей. В большинстве групп эта разница была статистически достоверной. Найдена также зависимость между характером оборонительного поведения и плодовитостью самок. Наибольшая плодовитость обнаружена у спокойных самок, наименьшая — у злобно-трусливых. Разница между этими группами статистически достоверна. Статистически достоверные различия (в первый год спаривания) обнаружены и между числом потомков спокойных самок по сравнению с числом щенят, рождающихся от злобных и трусливых самок. Обнаружена интересная зависимость между особенностями поведения и окраской. Наибольшее количество серебристого (зонарно окрашенного) волоса оказывается у лисиц с той или другой формой оборонительного поведения. Среди злобных лисиц оказался наименьший процент особей с низким содержанием серебристых волос. Среди лисиц, селекционируемых по спокойной форме поведения (такая селекция оказалась эффективной), встречались особи с аномалиями в строении мехового покрова. Поскольку количество серебра в мехе лисиц увеличивает ценность шкурки, делается понятным, почему в хозяйствах сохраняются злобные и трусливые особи (менее удобные при уходе за ними) и почему отбор лисиц по времени наступления у них течки, который ведется в звероводческой практике, не дает достаточного эффекта. Проводимое Беляевым исследование показывает значение изучения генетики поведения в селекции по хозяйственно полезным признакам в звероводческих хозяйствах, открывает новые пути подхода к проблеме одомашнивания животных и показывает роль поведения в формировании морфофизиологических особенностей популяции. Выше были изложены данные, показывающие роль действия генов на формирование поведения особи на разных уровнях ее индивидуального развития. Однако генотипические факторы, как это выяснено в настоящее время, оказывают свое влияние на поведение животных также посредством установления различных взаимоотношений между особями в отдельных сообществах. Одним из наиболее хорошо изученных путей, посредством которого генотип оказывает влияние на групповое поведение, является степень выраженности агрессии по отношению к особям своего вида. В каждом сообществе позвоночных животных в результате агрессивности к особям своего вида устанавливается поведенческая иерархия: одни особи оказываются «преобладающими», другие «подчиненными»; «подчиненные» особи боятся «преобладающих». Иерархическая система поведения сообщества зависит от многих факторов. Как правило, молодые особи оказываются подчиненными более старым. Самцы вне периода размножения оказываются преобладающими над самками. Однако в сезон размножения, как это показано на птицах, самки начинают преобладать над самцами. Самцы, стоящие на самой низкой ступени иерархии сообщества, не выбираются самками птиц для спаривания. Самки, которые стоят на низкой ступени иерархии сообщества, если они спариваются с агрессивными, преобладающими самцами, сами начинают занимать преобладающее положение в сообществе. Большую роль в том месте, которое займет данная особь в системе иерархии в сообществе, как показал ряд исследований, играют генотипические факторы. Это было отчетливо показано на мышах. Мыши различных инбредных линий имеют разную степень агрессивности, определяющей иерархию в сообществе этих животных. Так, например, из этих линий мышей мыши линии C57BL/10 (черные) имеют наибольшую тенденцию занимать преобладающее положение, затем идут мыши линии СЗН (зонарно-серые) и наиболее подчиненными оказываются мыши линии BALB (белые). Однако, несмотря на ясную роль генотипа, определяющую структуру сообщества у мышей, выяснилась и большая роль условий воспитания для степени агрессивности каждой мыши. Оказалось, что мыши легко подчиняемых линий при изолированном воспитании делаются более агрессивными и начинают подчинять мышей тех линий, которые воспитывались в сообществе. Однако если мышь из малоагрессивной линии воспитывается вместе с мышами агрессивной линии, ее агрессивность повышается. Помещенная затем в сообщество малоагрессивных мышей, она займет высокое место в иерархии подчинения. Большую роль в формировании иерархической структуры сообщества в семействе хищных оказывает генотип. Исследования, проведенные Павловским и Скоттом, показали, что степень выраженности преобладания у собак определяется генотипом и чрезвычайно различна среди разных пород. Это различие выступает в яркой степени начиная с 11-недельного возраста. Наиболее ясное преобладание (особенно самцов над самками) обнаружено у африканских охотничьих собак (бейсенджи) и фокстерьеров. У бигль и коккер-спаниелей это преобладание носит весьма стертый характер (рис. 4). Резко выраженная иерархия поведения в сообществе собак делает чрезвычайно затруднительным вступление в это сообщество новых особей. Слабо выраженная иерархия поведения у бигль и спаниелей является, по мнению авторов, результатом длительного искусственного отбора, при котором выбраковывались наиболее агрессивные особи, не дававшие возможности включения в стаю новых собак. Резкая выраженность иерархии в сообществе семейства собачьих наблюдается у диких видов, например у волков и у наименее одомашненных пород собак, и имеет большое биологическое значение в борьбе за территорию обитания. Так, в эскимосских поселках образуется несколько сообществ лаек. Щенки, как правило, могут безнаказанно ходить по всему поселку. Однако после наступления половой зрелости, когда они начинают спариваться, каждая собака примыкает к какому-либо сообществу и после этого загрызается, если заходит на территорию, занимаемую другим сообществом (Тинберген). У диких животных обычно образуется сообщество, состоящее из различного числа членов. Это может быть семейство или стадо, не связанное непосредственным родством. Одна из основных причин, определяющая образование таких сообществ, это защита занимаемой территории от проникновения на нее членов других сообществ. Число особей, входящих в сообщество, в значительной степени определяется генотипом, как это было показано в работе Леопольда на индюках. Оказалось, что дикие индейки образуют более мелкие стаи, чем особи гибридной популяции. Таким образом, генотипически обусловленная агрессивность по отношению к особям своего вида имеет биологическое значение. Во-первых, она способствует образованию физиологически изолированных групп биотипов, что является важнейшим условием видообразования. Во-вторых, создавая иерархическую систему поведения, агрессивность ставит наиболее слабых особей в наименее выгодные условия для размножения, что благоприятствует отрицательному отбору наименее приспособленных особей. И наконец, в-третьих, агрессивность по отношению к особям своего вида приводит, как это было в яркой форме показано К. Лоренцом на примере борьбы за существование, осуществляемой среди рыб коралловых рифов, к равномерному распределению особей одного и того же вида по всей территории обитания, что обусловливает ее наиболее рациональное использование. Таким образом, специфика действия генов сводится к тому, что они не только играют важнейшую роль в формировании поведения отдельной особи, но и в значительной степени определяют взаимоотношения животных в пределах отдельных сообществ, оказывая тем самым влияние на формирование структуры популяции и ход эволюционного процесса. Применение генетических методов для изучения физиологических механизмов, лежащих в основе патологических нарушений высшей нервной деятельности, ясно видно по интенсивному изучению во многих странах мира наследственных заболеваний нервной системы у животных. Наиболее широко изучаемой генетической моделью болезни нервной системы является экспериментальная эпилепсия грызунов. В 1907 г. была обнаружена мутация у кроликов породы венские белые, у которых эпилептические припадки развивались при действии различных неспецифических внешних раздражителей. Нахтсхейм путем инбридинга добился появления припадков у 75 % кроликов своей линии. Он пришел к выводу, что предрасположенность к судорожным припадкам определяется одним рецессивным геном. Однако на наследственное осуществление этой особенности поведения оказывают тормозящее влияние несколько модификаторов. Линия кроликов Нахтсхейма погибла во время второй мировой войны. В настоящее время выведена линия, в которой почти 100 % особей отвечают судорожными припадками в ответ на действие сильных звуковых раздражителей. Широкое распространение в качестве экспериментальной модели эпилепсии получили судорожные припадки мышей и крыс. Вит и Холл исследовали генетику судорожных припадков (так называемой аудиогенной или рефлекторной эпилепсии) у мышей, получаемых действием звукового раздражителя (обычно используется звук электрического звонка силой 100–120 Дб). Авторы производили скрещивание двух инбредных линий: С57 BL, у которых в ответ на звуковое раздражение судорожные припадки развиваются в 5 % случаев, с линией DBA, у которых припадки развиваются у 95 % особей. Они пришли к выводу о том, что повышенная предрасположенность к судорожным припадкам определяется одним доминантным геном. Однако последующие исследования, проведенные на тех же линиях мышей, не подтвердили монофакториальной картины исследования. Гинзбург и Старбук-Миллер в результате своих многолетних наследований приходят к выводу, что сходное фенотипическое выражение судорожных припадков, наблюдаемое в разных сублиниях мышей (С57 BL/6 и С57 BL/10, DBA/1 и DBA/2), имеет разную генетическую основу. Наиболее вероятно, что «чувствительные» к судорожным припадкам сублинии DBA имеют генетическую формулу ААВВ, а малочувствительные сублинии С57 BL имеют два рецессивных аллеля аавв. Каждый из этих генов расположен в разных аутосомах. Наиболее вероятное различие в степени доминирования и соотношения чувствительных и нечувствительных особей в F2 и обратных скрещиваниях объясняется различием каждой из использованных линий по ряду генов модификаторов. Физиологические исследования, проведенные параллельно с генетическим анализом, показали, что предрасположенность к эпилептическим припадкам у мышей связана с их общей предрасположенностью к действию стресс-факторов.[42] Особенно большое значение при этом, по-видимому, имеют окислительно-фосфорилирующие механизмы, которые различны у исследованных линий мышей. Фенотипическое появление и выражение судорожной готовности мышей находятся под большим влиянием различных внешних факторов. Ярким примером такого влияния является увеличение судорожной готовности мышей линии DBA/1 и их гибридов, полученных от скрещивания с мышами С57 BL при действии очень небольших доз радия. Мыши, которые с момента рождения в течение месяца подвергались хронической экспозиции гамма-лучей (общая доза 0,14 rad), оказались повышенно чувствительными к действию звукового раздражителя. Изменение общего фона радиации оказывает также влияние на чувствительность мышей к действию звукового раздражителя. С мая по октябрь 1957 г. у мышей линии DBA/1 в Fi, полученного от скрещивания с С57 BL, Старбук-Миллер обнаружила значительное увеличение числа судорожных припадков при звуковой экспозиции. По данным комиссии по атомной энергии, этот период совпал с повышением общего уровня радиации в Америке. Развитие судорожных припадков у крыс под влиянием звукового раздражителя обусловлено также генотипически. В популяции лабораторных крыс и линии крыс Уистар около 10–15 %особей дают судорожные припадки в ответ на звуковой раздражитель (звонок 100–120 Дб). В результате селекции нам удалось получить линию крыс, дающих судорожные припадки при звуковой экспозиции у 98–99 % особей. Несмотря на ясную генотипическую детерминированность этой патологической реакции крыс, точная картина ее наследования неясна. Опубликованные различными авторами данные противоречивы. Полученные нами с Л. Н. Молодкиной данные указывают лишь на очевидное неполное доминирование повышенной чувствительности крыс к действию звуковых раздражителей. При скрещивании крыс нашей линии с нечувствительными неселекционированными крысами и крысами линии Уистар в первом поколении оказалось чувствительных особей 93 (69,9 %), нечувствительных 40 (30,1 %). На основании характера расщепления в последующих поколениях пока трудно сделать вывод о количестве наследственных факторов, контролирующих развитие данной патологии. Однако в сложном комплексе реакции крыс на звуковой раздражитель удалось выделить более просто наследуемые особенности нервной деятельности. Таким свойством оказалось затяжное возбуждение. Оно выражается в том, что крыса после нескольких минут (по нашему стандарту 8) звуковой экспозиции, несмотря на выключение раздражителя, продолжает находиться в состоянии сильного двигательного возбуждения, длящегося иногда десятки минут (рис. 4). Это свойство в яркой форме было обнаружено у одного самца нашей чувствительной линии. В результате селекции и инбридинга эта особенность нервной деятельности была закреплена. Затяжное возбуждение оказалось рецессивным признаком по отношению к отсутствию этого функционального свойства нервной деятельности: все 68 чувствительных к звуковому раздражителю гибридов Fi, полученных от скрещивания крыс нашей линии с неселекционированными крысами и крысами линии Уистар, у которых это свойство отсутствует, оказались без затяжного возбуждения. При обратном скрещивании F1 с линейными крысами, обладающими затяжным возбуждением, из 93 чувствительных к звуковому раздражителю особей 70 оказались без затяжного возбуждения и 23 — с затяжным. Это расщепление согласуется с гипотезой о том, что затяжное возбуждение обусловливается двумя рецессивными генами. При этом ожидаемое расщепление должно быть 69,75:23,25. Однако при скрещивании между собой крыс с затяжным возбуждением, полученных от этого скрещивания, наряду с крысами, обладающими затяжным возбуждением, рождаются и особи без него. Это указывает на то, что анализируемый признак контролируется более сложно, чем двумя рецессивными генами, проявляющимися в 100 % случаев. Выведение высокочувствительной к звуковому раздражению линии крыс, обладающих затяжным возбуждением, явилось важнейшим шагом для возможности проведения патофизиологических исследований. У крыс нашей линии, помимо эпилептических припадков, развивается целый ряд патологий: важнейшие из них касаются сердечно-сосудистой системы — это смерть от кровоизлияний в головной мозг, изменение кровяного давления, функциональные нарушения (аритмии) сердечной деятельности и др. Основной причиной развития всех этих патологий является возбуждение мозга под влиянием действия звукового раздражителя. Генотипическая детерминированность порога возбуждения и силы защитно-тормозных процессов, с одной стороны, и функциональное состояние нервных центров, зависящее от ряда внешних факторов, — с другой, определяют картину развивающегося возбуждения. Однако, несмотря на множественность факторов, принимающих участие в развитии данной патологии, относительно простые отношения между процессом возбуждения и торможения определяют все многообразие наблюдаемых стадий этого процесса. Эти отношения, как было показано Савиновым, Крушинским, Флессом и Валлерштейн, очевидно, сводятся к тому, что возбуждение в течение времени действия звукового раздражителя растет, приближаясь к кривой, имеющей линейный характер, а тормозной процесс, ограничивающий это возбуждение, растет по экспоненте. Исследованиями, проводимыми по генетике поведения, установлен ряд фактов. Во-первых, показано, что многие акты поведения контролируются небольшим числом генов, наследуясь по законам Менделя. В результате комбинации независимо наследуемых актов поведения формируются более сложные, целостные в своем проявлении и выражении формы поведения, которые могут быть расчленены как генетическими, так и физиологическими методами. В то же время сложный морфофизиологический комплекс, как, например, одомашненность, может обусловливаться отбором небольшого (даже одного) числа генов, оказывающих плейотропный эффект на поведение и морфологические признаки. Во-вторых, генный контроль поведения осуществляется на самых различных уровнях организации. Найдены различия в поведении, которые контролируются генами, действующими на клеточном уровне, генами, контролирующими биохимические и физиологические процессы, лежащие в основе разных типов поведения, и, наконец, генами, действующими на поведенческом уровне, определяя тем самым разную структуру популяции. С выяснением той роли, которую играют генотипические факторы, контролирующие поведение в формировании отдельных изолированных микробиотипов в популяции, и установлением роли генотипических факторов в разных типах активности открывается новое направление в изучении роли генетики в эволюции и биогеоценологии. В-третьих, наследственное осуществление реакций поведения находится в чрезвычайно большой зависимости от индивидуального опыта. Сходные по своему внешнему проявлению акты поведения могут обусловливаться различными причинами. В одних случаях они формируются под ведущим влиянием врожденных факторов, в других — под ведущим влиянием индивидуального опыта. Трудность в разграничении без специального генотипического анализа наследственной и ненаследственной изменчивости поведения и возможность передачи путем подражания определенных традиций от поколения к поколению издавна создавали условия для необоснованных допущений прямого наследования индивидуально приобретенных навыков. В-четвертых, установлена генотипическая обусловленность патологических реакций нервной деятельности, сходных с болезнями человека. Обнаруживаются определенные генотипически обусловленные сдвиги в общих биохимических и физиологических системах, которые могут лежать в основе широкого круга патологических реакций организма. Это открывает возможность для изучения в модельных опытах на животных с определенным генотипом биохимических и физиологических механизмов, лежащих в основе развития ряда патологий, с которыми встречается клиника. Нет никакого сомнения в том, что сближение генетики и физиологии высшей нервной деятельности не только обогатит обе эти науки, но и окажет большое влияние на ряд других разделов биологии. Примечания:3 Опытные показатели шагомеров, полученные после инъекции; контрольные показатели шагомеров, полученные до инъекции. 4 Журн. общ. биологии. 1944. Т. 5, № 5. С. 261–283. 39 Актуальные вопросы современной генетики. М.: Изд-во МГУ, 1966. С. 281–301. 40 Павлов И. П. Лекции по физиологии. Л., 1952. 41 Несмотря на то что по степени силы наблюдается непрерывная изменчивость между исследованными особями, в приведенной таблице все собаки разделены на две альтернативные группы: слабые и сильные. 42 Под стресс-факторами понимаются различные неспецифические раздражители, которые приводят к глубоким нарушениям регуляторных механизмов организма. |
|
|||
|
Главная | Контакты | Прислать материал | Добавить в избранное | Сообщить об ошибке |
||||
|
|
||||